A Review of the Curculionoidea (Coleoptera) from European Eocene Ambers


1
Institute of Systematics and Ecology of Animals, Siberian Branch, Russian Academy of Sciences, Frunze Street 11, 630091 Novosibirsk, Russia
2
Biological Institute, Tomsk State University, Lenina Prospekt 36, 634050 Tomsk, Russia
Geosciences 2020, 10(1), 16; https://doi.org/10.3390/geosciences10010016
Submission received: 16 October 2019
/
Revised: 18 December 2019
/
Accepted: 23 December 2019
/
Published: 30 December 2019
(This article belongs to the Special Issue The Evolutionary History of the Coleoptera)
Abstract
:All 142 known species of Curculionoidea in Eocene amber are documented, including one species of Nemonychidae, 16 species of Anthribidae, six species of Belidae, 10 species of Rhynchitidae, 13 species of Brentidae, 70 species of Curcuionidae, two species of Platypodidae, and 24 species of Scolytidae. Oise amber has eight species, Baltic amber has 118 species, and Rovno amber has 16 species. Nine new genera and 18 new species are described from Baltic amber. Four new synonyms are noted: Palaeometrioxena Legalov, 2012, syn. nov. is synonymous with Archimetrioxena Voss, 1953; Paleopissodes weigangae Ulke, 1947, syn. nov. is synonymous with Electrotribus theryi Hustache, 1942; Electrotribus erectosquamata Rheinheimer, 2007, syn. nov. is synonymous with Succinostyphlus mroczkowskii Kuska, 1996; Protonaupactus Zherikhin, 1971, syn. nov. is synonymous with Paonaupactus Voss, 1953. Keys for Eocene amber Curculionoidea are given. There are the first records of Aedemonini and Camarotini, and genera Limalophus and Cenocephalus in Baltic amber.
1. Introduction
The Curculionoidea are one of the largest and most diverse groups of beetles, including more than 62,000 species [1] comprising 11 families [2,3]. They have a complex morphological structure [2,3,4,5,6,7], ecological confinement, and diverse trophic links [1], which makes them a convenient group for characterizing modern and fossil biocenoses. The oldest Curculionoidea are described from the Middle Jurassic of China [8] and the Middle–Upper Jurassic of Kazakhstan [2,8,9,10,11,12]. The major families appear in the Lower Cretaceous [2,13,14,15,16,17], while the major subfamilies occur in the Palaeogene [2,17].
The Eocene ambers of Europe with Curculionoidea include the Earliest Eocene Oise amber and Late Eocene Baltic and Rovno ambers. Oise amber Coleoptera were first studied by Kirejtshuk and Nel [18], while the first weevil from Oise amber was described in the Brentidae [19]. In 2019, a review of weevils of the families Brentidae and Curculionidae from Oise amber was published [20]. The first Curculionoidea from Baltic amber was Hylesinites electrinus Germar, 1813 (Scolytidae) [21]. The second species from Baltic amber was described by Motschulsky in 1857 [22]. Unfortunately, it is not possible to establish a more accurate systematic position of Erirhinoides cariniger Motschulsky, 1857 based on its brief description [22], and the collection where the holotype was stored was lost. A review of Baltic amber bark beetles was given by Schedl [23], which is still the only general work on Baltic amber bark beetles. Voss [24,25] and Zherikhin [26] described many weevils from Baltic amber in three articles [24,25,26]; later, 15 species were also described [27,28,29,30,31,32,33,34,35,36,37]. In 2012, the present author began a study of the Curculionoidea in Baltic amber [2,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53] and described 48 species. Data on Curculionoidea are also given in general works devoted to amber [54,55,56,57,58], catalogs [59,60], and descriptions in scientific collections [61,62,63]. The curculionoid fauna from Eocene Rovno amber represented by three families, despite recent work [64,65,66,67,68,69,70,71,72,73], remains insufficiently studied.
This review summarizes all previous information about Curculionoidea from Eocene amber, provides keys for currently described species from Eocene amber, comments on the identification of taxa, and also describes new taxa.
2. Materials and Methods
The Baltic amber mines are located along the Baltic Sea coast mostly at the Amber quarry of Yantarny (former Palmnicken) near Kaliningrad in the Kaliningrad Region (Russia). Baltic amber from the Prussian Formation dates 34–48 million years old [74]. The amber from this deposit was probably produced by Pinus succinifera, Cerdus, Pseudolarix (Pinaceae), Agathis (Araucariaceae), Sciadopitys (Sciadopityaceae) (Sadowski et al. 2016) [75,76,77,78,79], or other trees.
Rovno amber is found in the Rovno Region (Ukraine) [80]. The amber dates the Late Eocene (Priabonian) [80,81]. The plant producing this amber is possibly the same as Baltic amber.
The Oise amber deposit is located near the town of Creil at a place known as “Le Quesnoy” (Paris Basin, Creil, Oise, France). The age of this site was estimated as Lowermost Eocene (about 53 Ma) [82]. The amber-producing tree is Aulacoxylon sparnacense (Combretaceae or Caesalpiniaceae), which could be related to extant Terminalia L. (Combretaceae) or Fabaceae–Caesalpiniaceae [83].
The type specimens and material considered below are housed in the following collections: (ABCD) A. Bukejs’s collection, Daugavpils, Latvia; (ANSP) Department of Entomology, Academy of Natural Sciences, Philadelphia, United States of America (USA); (CAGB) A. Górski’s collection, Bielsko-Biala, Poland; (CCMCL) Centre de Conservation du musée des confluences, Lyon, France; (CVGM) V. Gusakov’s collection, Russia, Moscow; (CVIA) V. Alekseev’s collection, Kaliningrad, Russia; (EIW) Earth Institute, Warsaw, Poland; (FEH) Friedhelm Eichmann, Hannover, Germany; (FKCH) F. Kernegger’s collection, Hamburg, Germany, deposited in the Forschungsinstitut Senckenberg, Frankfurt am Main, Germany; (GMPB) Górnośląskie Muzeum Przyrodnicze w Bytomiu, Poland; (GPIH) Center of Natural History (formerly Geological–Paleontological Institute and Museum), Hamburg, Germany; (GPIHG) C. Gröhn’s collection (Glinde, Germany) deposited in the Center of Natural History (formerly Geological–Paleontological Institute and Museum), Hamburg, Germany; (HCH) Ch. and H. W. Hoffeins’s collection, Hamburg, Germany; (ISEA) A. Legalov’s fossil insects collection maintained at Institute of Systematics and Ecology of Animals of the Siberian Branch of the Russian Academy of Science, Novosibirsk, Russia; (KRAM) Kaliningrad Regional Amber Museum, Kaliningrad, Russia; (MAIG) Museum of Amber Inclusions, University of Gdańsk, Poland; (MLUH) Martin Luther University, Halle, Germany; (MNHN) Muséum national d’histoire naturelle, Paris, France; (MWOK) Museum of the World Ocean, Kaliningrad, Russia; (PACO) Poinar amber collection maintained at Oregon State University, Corvallis, OR, USA; (PIN) Borissiak Paleontological Institute of the Russian Academy of Sciences, Moscow, Russia; (SDEI) Senckenberg Deutsches Entomologisches Institut, Müncheberg, Germany; (SFNF) Senckenberg Forschungsinstitut und Naturmuseum, Frankfurt, Germany; (SIZK) Schmalhausen Institute of Zoology of the National Academy of Sciences of Ukraine, Kiev, Ukraine; (SMNK) Staatliches Museum für Naturkunde, Karlsruhe, Germany; (SMNS) Staatlichen Museums für Naturkunde Stuttgart, Germany; (ZMHB) Museum für Naturkunde, Leibniz-Institut für Evolutions-und Biodiversitätsforschung, Berlin, Germany; (ZMUC) Zoological Museum, University of Copenhagen, Denmark.
Descriptions and photographs were produced using Leica and Zeiss Stemi 2000-C stereomicroscopes. Photographs 1a–1d, 1f–1i, 2a–2i, 3a, 3d–3g, 4a–4h, 4j, 5a–5j, 6a–6c, 6i–6m, 7a–7f, 7h–7k, 8a–8k, 9a–9f, 10a–10i, 11a, 11d, 11e, 11h–11j, and 12a–12i were taken by the author, while photographs 1e, 3b, 3c, 3h, 4i, 6d–6g, 7g, 11b, 11c, 11f, 13c, and 13g were received from Ulrich Kotthoff and Eva Vinx (Germany: Hamburg), photographs 5k, 5l, and 6h were received from Didier Berthet (Lyon), photographs 7h, 8d, and 11g were received from Katarzyna Szczepaniak (Poland: Warsaw).
The spectra of laser-induced fluorescence (LIF) of some samples were examined with the assistance of N.A. Maslov (Khristianovich Institute of Theoretical and Applied Mechanics of the Siberian Branch of the RAS) to confirm their origin.
Taxa known from amber from references without author examination are included in square brackets whose identity should be clarified by further re-examination.
The systematics of studied taxa are from Legalov’s works [2,3,84,85,86,87,88]. The terminology of weevil morphological structures is based on Lawrence et al. [89].
Nomenclatural acts introduced in the present work are registered in ZooBank (www.zoobank.org) under LSID urn:lsid:zoobank.org:pub: AF69581E-33D5-4EEB-9B71-60945570065B.
3. Results
Superfamily Curculionoidea Latreille, 1802
Key to families of superfamily Curculionoidea in Eocene amber
1. Head ventrally with pregular sutures. Rostral pleurostomal sinus shallow. Rostrum reduced. Tibiae with denticles on outer margin………………………………………………………………………2
—Head ventrally lacking pregular sutures. Rostral pleurostomal sinus usually deep; as exception, it can be shallow (Nemonychidae, Attelabidae). Rostrum distinct. Tibiae lacking denticles on outer margin………………………………………………………………………………………………3
2. Pregular sutures parallel to exterior margin of hypostomal sinus. Tibiae with denticles on outer margin. Head narrower than pronotum. Tarsomere 1 shorter than tarsomeres 2–5 combined. Club with sutures…………………………………………………………………………………Scolytidae
—Pregular sutures continued anteriorly to hypostomal margin at bottom of hypostomal sinus. Tibiae with transverse carinae on outer margin. Head as wide as pronotum. Tarsomere 1 longer thantarsomeres 2–5 combined. Club without sutures..........................................................Platypodidae
3. Labrum free. Maxillary palpi elongate……………………………………………………………4
—Labrum fused to head……………………………………………………………………………….5
4. Rostral pleurostomal sinus shallow. Pygidium absent. All five ventrites completely free. Tibiae with spurs.……………………………………………………………………………Nemonychidae
—Rostral pleurostomal sinus deep. Pygidium present and exposed. Ventrites 1–4 fused to greater or lesser extent, if free than pygidium exposed. Tibiae lacking spurs…………Anthribidae
5. Antennae geniculate.…………………………………………………………………………………6
—Antennae straight……………………………………………………………………………………7
6. Trochanters extended. Ventrites 1 and 2 and ventrites 3–5 oriented in different planes. Tibiae without uncus. ……………....................................................................................Brentidae (Nanophyinae)
—Trochanters short. Ventrites 1–5 oriented in one plane. Tibiae with uncus, if without uncus than ventrites 1–5 oriented in one plane. ……………………………………………………Curculionidae
7. Protibiae with wide apical groove and thick hairs on inner surface at point of tarsal attachment. Antennal scrobes absent. All five ventrites free. Tarsomeres 1–3 bilobed…………………………………………………………………………………………………Belidae
—Protibiae simple. Antennal scrobes distinct. At least ventrites 1 and 2 fused. Tarsomere 3 bilobed…………………………………………………………………………………………………………8
8. Ventrites 1 and 2 fused and elongate, and ventrites 3 and 4 shortened....... ……………………………………………………………………………Brentidae (without Nanophyinae)
—Ventrites more or less of similar length..........................……………………………………………9
9. Claws free at base. Tibiae of females at apex simple or with small mucro. Internal edge of protibiae without teeth. Mandible with teeth at external edge.………………………… Rhynchitidae
—Claws fused at base. Tibiae of females at apex with large mucro. Internal edge of protibiae with fine teeth. Mandible without teeth at external edge. ………………………………… [Attelabidae]
Nemonychidae Bedel, 1882
Cimberidinae Gozis, 1882
Kuschelomacerini Riedel, 2010
Genus Kuschelomacer Riedel, 2010
Type species: Kuschelomacer kerneggeri Riedel, 2010
Kuschelomacer kerneggeri Riedel, 2010
Riedel [35]: 32–36, Figures 1–13 therein.
Locality. Baltic amber.
Remarks. The holotype of this species deposited in FKCH.
Anthribidae Billberg, 1820
Key to subfamilies of Anthribidae in Eocene amber
1. Elytra lacking striae. Tarsomere 2 simple. Transverse carina absent. Ventrites free..…………………………………………………………………………………………… [Urodontinae]
—Elytra striate. Tarsomeres 2 partially embracing tarsomere 3 laterally. Transverse carina usually distinct, subbasal or basal. Ventrites 1–4 fused to some extent.....................................................2
2. Antennae inserted dorsally between or next to lower portion of eyes........................Choraginae
—Antennae inserted laterally or ventrally on rostrum....................................................Anthribinae
[Urodontinae C. G. Thomson, 1859]
Remarks. This is an undescribed representative of the subfamily recorded from Baltic amber [62].
Anthribinae Billberg, 1820
Key to tribes of Anthribinae in Eocene amber
1. Eyes coarsely facetted.........................................................................................................Oiserhinini
—Eyes finely facetted................................................................................................................................2
2 Rostrum shorter or equal in width.……………………………………………………Zygaenodini
—Rostrum longer than wide.…...............................................................................................................3
3. Antennal scrobes dorso-lateral partially visible from above.…………………………Mecocerini
—Antennal scrobes ventral or lateral, not visible from above.…………………………......………4
4. Rostrum perpendicular. Underside of head angulate to rostrum in profile………[Tropiderini]
—Rostrum directing antero-ventrally. Underside of head continuous to rostrum and forming an arc in profile.………………………………………………………………………………….…Allandrini
Figure 1.
Habitus of Anthribinae from Baltic amber. (a,b) Pseudomecocerus alekseevi gen. et sp. nov., holotype, KRAM, no. 6328: body, dorsally (a); body, laterally (b). (c) Allandroides vossi, holotype, ISEA, no. BA2012/11, body, laterally. (d,f,i) Glaesotropis balticus sp. nov., holotype, FEH, no. 656: body, dorso-laterally (d); body, laterally (f); body, latero-ventrally (i). (e) Pseudomecorhis simulator, holotype, GPIH, no. 193, body, laterally; (g,h) G. martynovi, holotype, PIN, no. 964/1235: body, ventrally (g); body, dorsally (h). Scale bars: 1.0 mm. See Section 2 for names of collections.
Figure 1.
Habitus of Anthribinae from Baltic amber. (a,b) Pseudomecocerus alekseevi gen. et sp. nov., holotype, KRAM, no. 6328: body, dorsally (a); body, laterally (b). (c) Allandroides vossi, holotype, ISEA, no. BA2012/11, body, laterally. (d,f,i) Glaesotropis balticus sp. nov., holotype, FEH, no. 656: body, dorso-laterally (d); body, laterally (f); body, latero-ventrally (i). (e) Pseudomecorhis simulator, holotype, GPIH, no. 193, body, laterally; (g,h) G. martynovi, holotype, PIN, no. 964/1235: body, ventrally (g); body, dorsally (h). Scale bars: 1.0 mm. See Section 2 for names of collections.

Oiserhinini Legalov, Kirejtshuk et Nel, 2019
Genus Oiserhinus Legalov, Kirejtshuk et Nel, 2019
Type species: Oiserhinus insolitus Legalov, Kirejtshuk et Nel, 2019
Oiserhinus insolitus Legalov, Kirejtshuk et Nel, 2019
Legalov et al. [20]: 64, 67, Figure 1a–e therein, plate 17, Figures 1–4 therein.
Locality. Oise amber.
Remarks. This species is known only by the holotype (MNHN).
Mecocerini Lacordaire, 1865
Genus Pseudomecocerus gen. nov.
urn:lsid:zoobank.org:act:107F7197-3237-4BA2-9B05-B921848119D7
Type species: Pseudomecocerus alekseevi sp. nov.
Diagnosis. Rostrum shorter than pronotum, 1.6 times as long as wide in middle, widened in apical third. Forehead wider than rostrum base. Eyes rounded, convex, finely faceted. Antennae long, 11-segmented, inserted dorso-laterally near apical third of rostrum. Antennal club indistinct. Antennal scrobes foveiform. Transverse carina subbasal. Lateral carina reaches second third of pronotum. Pronotal declivity narrow. Tarsi not elongate. Tarsomeres 2 embracing tarsomere 3 laterally.
Etymology. The name is formed from the Latin “pseudo-” (false) and the generic name “Mecocerus”. Gender masculine.
Comparison. The new genus differs from the genus Acanthothorax Gaede, 1832 in the shorter antennae, not elongate tarsi, rostrum weaker widened at the apex, and narrower pronotal declivity.
Remarks. The tarsomere 2 embracing tarsomere 3 laterally, distinct transverse carina on the pronotum, and antennae inserted dorso-laterally indicate that the new genus belongs to Anthribidae. The new genus belongs to Mecocerini based on the antebasal transverse pronotal carina, foveiform dorso-lateral antennal scrobes and 11-segmented antennae.
urn:lsid:zoobank.org:act:E0ACC250-C5FE-4349-9072-FEF9C79510EF
Description. Male. Size. Body length (without rostrum) 4.9 mm, rostrum length 0.7 mm. Body black, covered with short, light colored decumbent hairs. Head about equal to rostrum. Rostrum straight, 0.7 times as long as pronotum, 1.2 times as long as wide at apex, 1.6 times as long as wide in middle, 1.5 times as long as wide at base, densely punctate, flattened, widened in apical third. Forehead slightly wider than rostrum base, weakly convex, densely punctate. Eyes large, rounded, sharply convex, finely faceted. Temples 0.5 times as long as eye. Vertex slightly convex. Antennae long, inserted dorso-laterally near apical third of rostrum. Scape 1.3 times as long as wide. Antennomeres: 3–11—elongate; 3—2.5 times as long as wide; 4—2.9 times as long as wide, equal in length, and 0.9 times as narrow as antennomere 3; 5—3.1 times as long as wide, 1.1 times as long as and equal in width to antennomere 4; 6—2.7 times as long as wide, 0.7 times as long as and 0.9 times as narrow as antennomere 5; 7—3.3 times as long as wide, 1.3 times as long as and equal in width to antennomere 6; 8—2.7 times as long as wide, 0.8 times as long as and equal in width to antennomere 7; 9—3.0 times as long as wide, 1.5 times as long as and 1.3 times as wide as antennomere 8; 10—equal in length and width, 0.3 times as long as and equal in width to antennomere 9; 11—1.5 times as long as wide, 1.5 times as long as and equal in width to antennomere 10. Antennal club indistinct. Pronotum bell-shaped, 1.3 times as long as wide apically, 0.9 times as long as wide medially and basally. Disc coarsely punctate, slightly convex. Transverse carina subbasal. Lateral carina reaches second third of pronotum. Posterior angular carina complete. Pronotal declivity 0.3 times as long as pronotum. Mesonotum. Scutellum 0.9 times as long as wide. Elytra elongate, 2.2 times as long as wide at base, 1.9 times as long as wide in middle, 2.5 times as long as wide in apical quarter. Humeri weakly smoothed. Interstriae wide, 2.0–3.5 times as long as with of striae, slightly flattened, finely and intensely punctate. Prosternum punctate. Legs. Tibiae elongate, subparallel. Tarsi elongate. Tarsomere: 1—elongate, conical; 2—wide, conical, bilobed, covers base of tarsomere 3; 3—bilobed, narrower than tarsomere 2; 5—elongate.
Material examined. Holotype (KRAM), no. 6328.
Etymology. The epithet of this new species is dedicated to Vitalii I. Alekseev (Kaliningrad Regional Amber Museum) who contributed to the studies of the Baltic amber Coleoptera.
Locality. Baltic amber.
Allandrini Pierce, 1930
Key to genera of Allandrini in Baltic amber
1. Antennae equal or longer than body..........................................................................Pseudomecorhis
—Antennae reaching middle of pronotum.........................................................................Allandroides
Genus Pseudomecorhis Voss, 1953
Type species: Pseudomecorhis simulator Voss, 1953
Key to species of genus Pseudomecorhis in Baltic amber
1. Rostrum long and thin. Body (without rostrum) larger (10.0 mm). Temples shorter than eyes.....................................................................................................................................................P. simulator
—Rostrum short and thick. Body (without rostrum) smaller (6.0 mm). Temples about 1.5 times as long as length of eye..........................................................................................................................P. orlovi
Pseudomecorhis orlovi Zherikhin, 1971
Zherikhin [26]: 199, 202, 203, Figure 1 therein.
Locality. Baltic amber.
Remarks. The holotype should be deposited in PIN; however, it was not found there.
Pseudomecorhis simulator Voss, 1953 (Figure 1e)
Voss [24]: 122–123, Figure 1 therein.
Locality. Baltic amber.
Remarks. This species is known only by the holotype (GPIH).
Genus Allandroides Legalov, 2015
Type species: Allandroides vossi Legalov, 2015
Allandroides vossi Legalov, 2015 (Figure 1c)
Legalov [2]: 1452–1453, 1454, Figure 1b therein, plate 1, Figure 2 therein.
Locality. Baltic amber.
Remarks. The holotype of this species deposited in ISEA and specimen in MWOK no.7657/833.
[Tropiderini Lacordaire, 1865]
Remarks. The genus Tropideres Schoenherr, 1823 was recorded from Baltic amber [61].
Zygaenodini Lacordaire, 1865
Genus Glaesotropis Gratshev et Zherikhin, 1995
Type species: Glaesotropis weitschati Gratshev et Zherikhin, 1995
Key to subgenera of genus Glaesotropis in Eocene amber
1. Forehead narrower than longitudinal eye diameter...........................................Pseudoglaesotropis
—Forehead wider than longitudinal eye diameter...............................................................................2
2. Antennomere 2 weakly narrower than antennomere 3. Antennomeres 1–3 subequal in length........................................................................................................................... Glaesotropis sensu stricto
—Antennomere 3 distinctly narrower and more or less shorter than antennomere 2................................................................................................................................................... Electranthribus
Subgenus Pseudoglaesotropis Legalov, 2012, stat. nov.
Type species: Pseudoglaesotropis martynovi Legalov, 2012
Key to species of subgenus Pseudoglaesotropis in Baltic amber
1. Elytral striae absent. Rostrum narrowed to eye. Antennomeres 3 and 4 elongate................................................................................................................................ G. balticus sp. nov.
—Elytral striae regular. Rostrum subparallel. Antennomeres 3 and 4 short................ G. martynovi
urn:lsid:zoobank.org:act:FFF9CEF9-C27E-49EF-BBF2-3EC6EEE5FD65
Description. Male. Size. Body length (without rostrum) 2.2 mm, rostrum length 0.5 mm. Body black, covered with sparse, light colored appressed hairs. Head. Rostrum short, almost straight, 0.3 times as long as pronotum, 1.2 times as long as wide at apex, equal in width in middle, 1.8 times as long as wide at base, densely punctate, flattened, widened in apical third. Forehead 0.7 times as long as wide at rostrum base, 0.9 times as narrow as longitudinal eye diameter, weakly flattened, densely punctate. Eyes large, rounded, convex, finely faceted. Temples very short. Vertex slightly convex. Antennae long, inserted laterally near middle of rostrum. Scape 2.0 times as long as wide. Antennomeres: 2–8—conical; 2—2.6 times as long as wide, 1.6 times as long as and 1.3 times as wide as scape; 3—3.4 times as long as wide, 0.9 times as long as and 0.7 times as narrow as antennomere 2; 4—2.3 times as long as wide, 0.7 times as long as and equal in width to antennomere 3; 5—1.8 times as long as wide, 0.9 times as long as and 1.1 times as wide as antennomere 4; 6—1.6 times as long as wide, equal in length and 1.1 times as wide as antennomere 5; 7—equal in length and width, 0.7 times as long as and 1.1 times as wide as antennomere 6; 8—equal in length and width, 1.2 times as long as and 1.2 times as wide as antennomere 7; antennal club loose, 0.5 times as long as antennomeres 2–8 combined; 9—equal in length and width, 1.7 times as long as and 1.7 times as wide as antennomere 8; 10—0.5 times as long as wide, 0.7 times as long as and 1.4 times as wide as antennomere 9; 11—0.8 times as long as wide, 1.5 times as long as and 0.9 times as narrow as antennomere 10. Pronotum bell-shaped, coarsely punctate, slightly convex. Transverse carina subbasal. Lateral carina absent. Posterior angular carina complete. Pronotal declivity about 0.1 times as long as pronotum. Mesonotum. Scutellum small. Elytra quite wide, 1.7 times as long as pronotum. Humeri weakly smoothed. Elytral striae absent. Interstriae convex. Thorax. Prosternum coarsely punctate. Precoxal part of prosternum equal to procoxa length. Procoxal cavities rounded, separated. Postcoxal part of prosternum 0.5 times as long as procoxa length. Mesocoxal cavities rounded and separated. Metaventrite 1.3 times as long as length of mesocoxa, convex, densely punctate. Metepisternum 5.8 times as long as wide in middle. Metacoxal cavities dilated. Abdomen convex, weakly flattened in middle. Ventrite 1, 0.7 times as long as metacoxae. Ventrite 2 equal to ventrite 1. Ventrite 3, 0.7 times as long as ventrite 2. Ventrite 4, 0.8 times as long as ventrite 3. Ventrite 5 about 1.8 times as long as ventrite 4. Pygidium exposed. Legs elongate. Procoxae spherical. Femora clavate, without tooth. Profemora 2.5 times as long as wide in middle. Mesofemora 2.6 times as long as wide. Metafemora about 2.0 times as long as wide. Tibiae almost straight, weakly flattened, without mucro. Metatibia 5.2 times as long as wide in middle. Tarsi elongate, shorter than tibiae, with pulvilli on underside. Tarsomeres: 1—elongate, conical; 2—wide, conical, bilobed, covers base of tarsomere 3; 3—bilobed, narrower than second; 5—elongate. Claws large, diverging, free, with teeth. Metatarsomeres: 1—1.3 times as long as wide; 2—1.3 times as long as wide, 1.1 times as long as and 1.1 times as wide as tarsomere 1; 3—0.8 times as long as wide, 0.4 times as long as and 0.7 times as narrow as tarsomere 2; 5—2.7 times as long as wide, 2.7 times as long as and 0.8 times as narrow as tarsomere 3.
Material examined. Holotype (FEH), no. 656.
Comparison. The new genus differs from G. (P.) martynovi in the elytra lacking elytral striae, rostrum narrowed to eye and elongate antennomere 3 and 4.
Etymology. The epithet of this new species is formed from the name of the Baltic Sea, on the coast where the amber sample was collected.
Locality. Baltic amber.
Pseudoglaesotropis martynovi Legalov, 2012
Legalov [39]: 263, 265, Figure 1a,b therein, pl. 8, Figure 1 therein.
Locality. Baltic amber.
Remarks. This species is known only by the holotype (PIN).
Subgenus Glaesotropis sensu stricto
Key to species of subgenus Glaesotropis sensu stricto in Eocene amber
1. Rostrum parallel-sided............................................................................................................G. minor
—Rostrum narrowing in the basal one-third........................................................................................2
2. Antennomeres 1 and 2 shorter than antennomeres 3 and 4 combined. Dorsal side of rostrum located in plane of forehead......................................................................................................G. diadiasashai
—Antennomeres 1 and 2 longer than antennomeres 3 and 4 combined. Dorsal side of rostrum separated by depression from forehead........................................................................................................3
3. Body 4.9–5.1 mm long. External elytral edge strongly concave at metacoxa. Forehead convex...............................................................................................................................................G. weitschati
—Body 2.6–3.8 mm long. External elytral edge slightly concave at metacoxa. Forehead flat........4
4. Relatively large (3.8 mm long). Body elongate. Forehead slightly convex.................G. gusakovi
—Relatively small (2.6–3.3 mm long). Body relatively wide. Forehead flat...............G. succiniferus
Figure 2.
Habitus of Anthribinae from Baltic amber. (a) Glaesotropis minor, holotype, PIN, no. 964/801: body, laterally. (b) G. minor, paratype, PIN, no. 363/109, body, laterally. (c) G. alleni, holotype, ISEA, no. BA2012/19, body, laterally. (d) G. gratshevi, holotype, ISEA, no. BA2014/2, body, laterally. (e) G. zherikhini, holotype, ISEA, no. BA2011/2, body, laterally. (f) G. weitschati, specimen, CVGM, no. 027C10, body, laterally. (g) G. gusakovi, holotype, CVGM, no. 026C12, body, laterally. (h) G. succiniferus, holotype, CVGM, no. 028C13, body, laterally. (i) Pseudomecocerus alekseevi gen. et sp. nov., holotype, KRAM, no. 6328, rostrum and head, dorsally. Scale bars: 1.0 mm. See Section 2 for names of collections.
Figure 2.
Habitus of Anthribinae from Baltic amber. (a) Glaesotropis minor, holotype, PIN, no. 964/801: body, laterally. (b) G. minor, paratype, PIN, no. 363/109, body, laterally. (c) G. alleni, holotype, ISEA, no. BA2012/19, body, laterally. (d) G. gratshevi, holotype, ISEA, no. BA2014/2, body, laterally. (e) G. zherikhini, holotype, ISEA, no. BA2011/2, body, laterally. (f) G. weitschati, specimen, CVGM, no. 027C10, body, laterally. (g) G. gusakovi, holotype, CVGM, no. 026C12, body, laterally. (h) G. succiniferus, holotype, CVGM, no. 028C13, body, laterally. (i) Pseudomecocerus alekseevi gen. et sp. nov., holotype, KRAM, no. 6328, rostrum and head, dorsally. Scale bars: 1.0 mm. See Section 2 for names of collections.

Glaesotropis (Glaesotropis) diadiasashai Gratshev et Perkovsky, 2008
Gratshev, Perkovsky [64]: 60–61, Figure 1 therein.
Locality. Rovno amber.
Remarks. This species is known only by the holotype (SIZK).
Legalov [2]: 1455–1456, Figure 1c therein, plate 1, Figure 3 therein.
Locality. Baltic amber.
Remarks. This species is known by the holotype (CVGM) and the specimen (MWOK) no. 7657/95.
Gratshev, Zherikhin [33]: 153, 155, Figures 6–10 therein, pl. I, Figure 2 therein.
Locality. Baltic amber.
Remarks. Holotype is deposited in the PIN.
Legalov [2]: 1455, 1456–1457, Figure 1c therein, plate 1, Figure 4 therein.
Locality. Baltic amber.
Remarks. This species is known by the holotype (CVGM) and paratype (ISEA).
Gratshev, Zherikhin [33]: 151, 153, Figures 1–5 therein, pl. I, Figure 1 therein.
Locality. Baltic amber.
Remarks. This species is known by the holotype (GPIH) and specimens (CVGM) no. 027C10, (MWOK) nos. 57/153 and 1155/218.
Subgenus Electranthribus Legalov, 2013
Type species: Electranthribus zherikhini Legalov, 2013
Key to species of subgenus Electranthribus in Baltic amber
1. Precoxal part of prosternum strongly elongate, 8.1 times as long as procoxa. Pronotal declivity relatively long. Relatively small (1.9 mm long)..........................................................G. gratshevi
—Precoxal part of prosternum only slightly elongate, subequal in length to procoxa. Pronotal declivity relatively short. Relatively large (2.2–2.8 mm long)...................................................................2
2. Posterior angular carinula incomplete. Antennomere 2 2.0 times as long as antennomere 3................................................................................................................................................................G. alleni
—Posterior angular carinula complete. Antennomere 2 1.5 times as long as antennomere 3.........................................................................................................................................................G. zherikhini
Legalov [2]: 1457, 1457–1458, Figure 1e therein, plate 1, Figure 5 therein.
Locality. Baltic amber.
Remarks. This species is known only by the holotype (ISEA).
Legalov [2]: 1457, 1458, Figure 1f therein, plate 2, Figure 2 therein.
Locality. Baltic amber.
Remarks. This species is known only by the holotype (ISEA).
Legalov [41]: 61, Figures 1–4 therein (Electranthribus).
Locality. Baltic amber.
Remarks. This species is known only by the holotype (ISEA).
Choraginae W. Kirby, 1819
Valenfriesiini Alonso-Zarazaga et Lyal, 1999
Genus Eduardoxenus Legalov, Nazarenko et Perkovsky, 2018
Type species: Eduardoxenus unicus Legalov, Nazarenko et Perkovsky, 2018
Eduardoxenus unicus Legalov, Nazarenko et Perkovsky, 2018
Legalov et al. [70]: 209–210, Figures 1–4 therein.
Locality. Rovno amber.
Remarks. This species is known only by the holotype (SIZK).
Belidae Schoenherr, 1826
Oxycoryninae Schoenherr, 1840
Key to tribes of Oxycoryninae in Baltic amber
1. Head behind eyes without constriction. Pronotum without carinae on disc. Rostrum in males without ventral long setae........................................................................................................Oxycraspedini
—Head behind eyes with more or less distinct constriction. Pronotum with serrated lateral carinae and three carinae on disc. Rostrum in males ventrally often with long setae.........Metrioxenini
Oxycraspedini Marvaldi et Oberprieler, 2006
Genus Oxycraspedus Kuschel, 1955.
Type species: Oxycraspedus poinari Legalov, 2016.
Oxycraspedus poinari Legalov, 2016 (Figure 3a)
Legalov [44]: 58–60, Figure 1 therein.
Locality. Baltic amber.
Remarks. The holotype of this species deposited in ISEA.
Metrioxenini Voss, 1953
Key to subtribes of Metrioxenini in Baltic amber
1. Ventrite 1 little longer than ventrite 2. Body large (2.2–2.6 mm in length)......................................................................................................................................Zherichinixenina
—Ventrite 1 strongly elongate. Body small (3.5–4.3 mm in length)..............................Metrioxenina
Metrioxenina Voss, 1953
Genus Archimetrioxena Voss, 1953
Type species: Archimetrioxena electrica Voss, 1953
= Palaeometrioxena Legalov, 2012, syn. nov.
Type species: Palaeometrioxena zherikhini Legalov, 2012
Remarks. The study of the holotype of Archimetrioxena electrica showed that this species also has blunt teeth along the edges of the pronotum.
Key to species of genus Archimetrioxena in Baltic amber
1. Forehead with small horn-like tubercles on either side of eyes. Pronotum densely punctate, almost matte....................................................................................................................................... A. electrica
—Forehead simple. Pronotum sparsely punctate, lustrous............................................. A. zherikhini
Figure 3.
Habitus of Belidae and Rhynchitidae from Baltic amber. (a) Oxycraspedus poinari, holotype, ISEA, no. BA2011/1, body, dorsally. (b,c) Archimetrioxena electrica, holotype, GPIH, no. 194: body, dorso-laterally (b); body, laterally (c). (d) A. electrica, specimen, ISEA, no. 2012/18, body, dorsally. (e) A. zherikhini, holotype, PIN. no. 964/1236, body, dorsally. (f) Succinometrioxena poinari, holotype, ISEA, no. BA2015/2, body, laterally. (g) S. bachofeni, holotype, ISEA, no. BA2012/6, body, dorsally. (h) Baltoconapium anderseni, holotype, ZMUC, no. 959, body, ventrally. (i) Baltocar succinicus, holotype, GPIH, no. 195, body, laterally, in front. (j) Palaeotanaos oisensis, specimen, MNHN, no. PA533, body, laterally. Scale bars: 1.0 mm. See Section 2 for names of collections.
Figure 3.
Habitus of Belidae and Rhynchitidae from Baltic amber. (a) Oxycraspedus poinari, holotype, ISEA, no. BA2011/1, body, dorsally. (b,c) Archimetrioxena electrica, holotype, GPIH, no. 194: body, dorso-laterally (b); body, laterally (c). (d) A. electrica, specimen, ISEA, no. 2012/18, body, dorsally. (e) A. zherikhini, holotype, PIN. no. 964/1236, body, dorsally. (f) Succinometrioxena poinari, holotype, ISEA, no. BA2015/2, body, laterally. (g) S. bachofeni, holotype, ISEA, no. BA2012/6, body, dorsally. (h) Baltoconapium anderseni, holotype, ZMUC, no. 959, body, ventrally. (i) Baltocar succinicus, holotype, GPIH, no. 195, body, laterally, in front. (j) Palaeotanaos oisensis, specimen, MNHN, no. PA533, body, laterally. Scale bars: 1.0 mm. See Section 2 for names of collections.

Archimetrioxena electrica Voss, 1953 (Figure 3b,d)
Voss [24]: 124–125, Figures 2 and 3 therein.
Locality. Baltic amber.
Remarks. This species is known by the holotype (GPIH) and specimen (ISEA no. 2012/18).
Archimetrioxena zherikhini (Legalov, 2012), comb. nov. (Figure 3e)
Palaeometrioxena zherikhini Legalov, 2012
Legalov [39]: 268, 270, Figure 2a,b therein, pl. 9, Figure 1 therein.
Locality. Baltic amber.
Remarks. This species is known by the holotype (PIN) and specimen (incl. GPIH, no. AKBS00111a-Curculionidae).
Zherichinixenina Legalov, 2009
Genus Succinometrioxena Legalov, 2012
Type species: Succinometrioxena poinari Legalov, 2012
Key to species of genus Succinometrioxena in Baltic amber
1. Forehead with horn-like tubercles on either side of eyes...................................................S. poinari
—Forehead without horn-like tubercles on either side of eyes...........................................................2
2. Body (without rostrum) smaller (3.5 mm). Elytra wider, with small punctation.....S. bachofeni
—Body (without rostrum) larger (6.3 mm). Elytra narrower, with large punctation.....S. attenuata
Succinometrioxena attenuata Legalov et Poinar, in lit.
Legalov, Poinar [53], in litteris.
Locality. Baltic amber.
Remarks. This species is known only by the holotype (PACO).
Succinometrioxena bachofeni Legalov, 2013 (Figure 3g)
Legalov [41]: 62–63, Figures 5–8 therein.
Locality. Baltic amber.
Remarks. This species is known only by the holotype (ISEA).
Succinometrioxena poinari Legalov, 2012 (Figure 3f)
Legalov [38]: 215–216, Figures 1–5 therein.
Locality. Baltic amber.
Remarks. Holotype is kept in ISEA.
Rhynchitidae Gistel, 1848
Key to subfamilies of Rhynchitidae in Eocene amber
1. Tarsal claws strongly divergent, lacking teeth...........................................................Sayrevilleinae
—Tarsal claws slightly divergent, usually with teeth...................................................Rhynchitinae
Sayrevilleinae Legalov, 2003
Sanyrevilleini Legalov, 2003
Genus Baltocar Kuschel, 1992
Type species: Car succinicus Voss, 1953
Remarks. The key to species is based on the key from Riedel et al. [36].
Key to species of genus Baltocar in Baltic amber
1. Elytral striae irregular. Rostrum shorter or subequal to pronotum................................................2
—Elytral striae regular. Rostrum longer or subequal to pronotum...................................................3
2. Body covered with dense setae. Rostrum slightly curved, subequal to pronotum....B. convexus
—Body without distinct pubescence. Rostrum distinctly curved, shorter than pronotum......................................................................................................................................... B. subnudus
3. Pronotal disc coarsely punctate. Setae widened........................................................... B. succinicus
—Pronotal disc transversely irregularly rugose. Setae thin................................................................4
4. Pronotum 1.4 times as long as wide, with subparallel sides............................................ B. groehni
—Pronotum 1.2 times as long as wide, with weak rounded from base to apex sides.............................................................................................................................................B. hoffeinsorum
Baltocar convexus Legalov, 2015 (Figure 4b)
Legalov [2]: 1499, 1501, Figure 5d therein, plate 7, Figure 1 therein.
Locality. Baltic amber.
Remarks. The holotype of this species deposited in ISEA.
Baltocar groehni Riedel, 2012
Riedel et al. [36]: 780–782, Figures 16–22 therein.
Locality. Baltic amber.
Remarks. This species is known by the holotype (GPIHG).
Baltocar hoffeinsorum Riedel, 2012 (Figure 4a)
Riedel et al. [36]: 782, 785–786, Figures 23–48, 65, and 66 therein.
Locality. Baltic amber.
Remarks. This species is known by the holotype (HCH) and three paratypes (GPIHG, SMNK) [36] and a specimen (ISEA) [2].
Baltocar subnudus Riedel, 2012
Riedel et al. [36]: 786, 789, Figures 49–58 therein.
Locality. Baltic amber.
Remarks. This species is known by the holotype (SDEI) and the paratype (GPIHG) [36].
Baltocar succinicus (Voss, 1953) (Figure 3i)
Voss [24]: 125–126, Figure 4 therein (Car).
Locality. Baltic amber.
Remarks. This species is known by the holotype (GPIH).
Rhynchitinae Gistel, 1848
Rhynchititae Gistel, 1848
Figure 4.
Habitus of Rhynchitidae and Brentidae from Baltic amber. (a) Baltocar hoffeinsorum, specimen, ISEA, no. 2012/14, body, laterally. (b) B. convexus, holotype, ISEA, no. BA2013/3, body, laterally. (c) Electrauletes unicus, holotype, ISEA, no. BA2014/1, body, laterally. (d) Succinorhynchites alberti, holotype, ISEA, no. BA2012/7, body, laterally. (e) Eocenorhynchites vossi, holotype, PIN. no. 964/1233, body, laterally. (f) Baltocyba electrinus, holotype, ISEA, no. BA2017/9, body, laterally. (g,h) Archinvolvulus liquidus, holotype, ZMUC, no. 957: fore-body, laterally (g); body, dorsally (h). (i) Baltoapion subdiscedens, holotype, GPIH, no. 196, body, laterally; (j) B. gusakovi, holotype, CVGM, no. 023C96, body, laterally. Scale bars: 1.0 mm. See Section 2 for names of collections.
Figure 4.
Habitus of Rhynchitidae and Brentidae from Baltic amber. (a) Baltocar hoffeinsorum, specimen, ISEA, no. 2012/14, body, laterally. (b) B. convexus, holotype, ISEA, no. BA2013/3, body, laterally. (c) Electrauletes unicus, holotype, ISEA, no. BA2014/1, body, laterally. (d) Succinorhynchites alberti, holotype, ISEA, no. BA2012/7, body, laterally. (e) Eocenorhynchites vossi, holotype, PIN. no. 964/1233, body, laterally. (f) Baltocyba electrinus, holotype, ISEA, no. BA2017/9, body, laterally. (g,h) Archinvolvulus liquidus, holotype, ZMUC, no. 957: fore-body, laterally (g); body, dorsally (h). (i) Baltoapion subdiscedens, holotype, GPIH, no. 196, body, laterally; (j) B. gusakovi, holotype, CVGM, no. 023C96, body, laterally. Scale bars: 1.0 mm. See Section 2 for names of collections.

Key to tribes of Rhynchititae in Eocene amber
1. Rounded apex of elytra when wings closed. Elytral striae indistinct or absent.…Auletini
—Apices of elytra separately rounded. Elytral striae regular………..……………Rhynchitini
Auletini Desbrochers des Loges, 1908
Key to subtribes of Auletini in Eocene amber
1. Tarsal claws lacking teeth.……………………………………………………………… Auletina
—Tarsal claws with teeth.…………………………………………………………….………..……2
2. Tibia lacking costate dorsal margin.…......................................................... Pseudomesauletina
—Tibia with costate dorsal margin.……………………………………..………… Pseudauletina
Auletina Desbrochers des Loges, 1908
Genus Electrauletes Legalov, 2015
Type species: Electrauletes unicus Legalov, 2015
Electrauletes unicus Legalov, 2015 (Figure 4c)
Legalov [2]: 1501–1502, Figure 5e therein, plate 7, Figure 3 therein.
Locality. Baltic amber.
Remarks. This species is known only by the holotype (ISEA).
Pseudauletina Voss, 1933
Genus Eoropseudauletes Kania et Legalov, 2019
Type species: Eoropseudauletes plucinskii Kania et Legalov, 2019
Eoropseudauletes plucinskii Kania et Legalov, 2019
Kania, Legalov [52]: 59, 61, Figures 1 and 2, pl. 6 therein.
Locality. Baltic amber.
Remarks. This species is known only by the holotype (GMPB).
Pseudomesauletina Legalov, 2003
Genus Pseudomesauletes Legalov, 2001
Type species: Auletes uniformis Roelofs, 1874
Subgenus Pseudomesauletes sensu stricto
Pseudomesauletes (Pseudomesauletes) groehni Bukejs et Legalov, 2019
Bukejs, Legalov [73], 169–170, Figure 1 therein.
Locality. Rovno amber.
Remarks. This species is known only by the holotype (GPIHG).
Rhynchitini Gistel, 1848
Key to subtribes of Rhynchitini in Baltic amber
1. Elytra lacking scutellar striole.………………………………………………………[Rhynchitina]
—Elytra with scutellar striole.…………………………………………………………………………2
2. Ventrite 1 narrower than ventrite 2. Ventrite 3 slightly narrower than ventrite 2......................................................................................................................................................Temnocerina
—Ventrites 1 and 2 almost equal in width. Ventrite 3 much narrower than ventrite 2....................................................................................................................................................Perrhynchitina
Temnocerina Legalov, 203
Genus Eocenorhynchites Legalov, 2012
Type species: Eocenorhynchites vossi Legalov, 2012
Eocenorhynchites vossi Legalov, 2012 (Figure 4e)
Legalov [39]: 266–267, Figure 1c,d therein, pl. 8, Figure 2 therein.
Locality. Baltic amber.
Remarks. This species is known only by the holotype (PIN).
Perrhynchitina Legalov, 2003
Genus Succinorhynchites Legalov, 2013
Type species: Succinorhynchites alberti Legalov, 2013
Succinorhynchites alberti Legalov, 2013 (Figure 4d)
Legalov [41]: 68–70, Figures 17 and 18 therein.
Locality. Baltic amber.
Remarks. This species is known only by the holotype (ISEA).
[Rhynchitina Gistel, 1848]
[Attelabidae Billberg, 1820]
Remarks. A tube rolled by a representative of this family was discovered in Baltic amber.
Brentidae Billberg, 1820
Key to subfamilies of Brentidae in Eocene amber
1. Antennae straight. Antennal scrobes deep. Scutellum visible. Base of elytra lacking crenulate carina..................................................................................................................................................Apioninae
—Antennae geniculate. Antennal scrobes shallow. Scutellum concealed. Base of elytra with crenulate carina............................................................................................................................Nanophyinae
Apioninae Schoenherr, 1823
Key to supertribes of Apioninae in Eocene amber
1. Elytral stria 10 distinct, merges with stria 9 near metacoxa.......……................... Rhadinocybitae
—Elytral stria 10 absent............................................................................................................................2
2. Procoxal cavities contiguous. Trochanters short......................................................Palaeotanaitae
—Procoxal cavities separated. Trochanters long..................................................................................3
3. Pronotal vestiture centrifugal, in which hairs directed away from pronotal midline basally, more or less parallel to midline on sides and perpendicular to apical edge apically......................................................................................................................................... Aspidapiitae
—Pronotal vestiture centripetal, in which hairs directed to pronotal midline............................................................................................................................................... Apionitae
Rhadinocybitae Alonso-Zarazaga, 1992
Key to tribes of Rhadinocybitae in Baltic amber
1. Antennomere 1 almost as long as antennomeres 2 and 3 combined. Greatest width of pronotum near middle. Elytral striae with sharp edges...........................................................Notapionini
—Antennomere 1 being slightly larger than antennomeres 2–5 combined. Greatest width of pronotum in apical third. Elytral striae lacking sharp edges...............................................Rhadinocybini
Rhadinocybini Alonso-Zarazaga, 1992
Genus Baltocyba Legalov, 2018
Type species: Baltocyba electrinus Legalov, 2018
Baltocyba electrinus Legalov, 2018 (Figure 4f)
Legalov [46]: 162–164, Figure 1 therein.
Locality. Baltic amber.
Remarks. This species is known only by the holotype (ISEA).
Notapionini Zimmerman, 1994
Genus Archinvolvulus Voss, 1972
Type species: Involvulus liquidus Voss, 1972
Archinvolvulus liquidus (Voss, 1972) (Figure 4g,h)
Voss [25]: 170–171, Figure 2 therein (Involvulus).
Locality. Baltic amber.
Remarks. This species is known only by the holotype (ZMUC).
Palaeotanaitae Legalov, Kirejtshuk et Nel, 2019
Palaeotanaini Legalov, Kirejtshuk et Nel, 2019
Genus Palaeotanaos Kirejtshuk, Legalov et Nel, 2015
Type species: Palaeotanaos oisensis Kirejtshuk, Legalov et Nel, 2015
Palaeotanaos oisensis Kirejtshuk, Legalov et Nel, 2015 (Figure 3j)
Kirejtshuk et al. [19]: 1437, 1439, 1440, Figures 1 and 2 therein.
Locality. Oise amber.
Remarks. This is a common species in Oise amber [20].
Aspidapiitae Alonso-Zarazaga, 1990
Key to tribes of Aspidapiitae in Baltic amber
1. Scutellum protrudes above elytra, distinctly elongate..............................……………Aspidapiini
—Scutellum flat, weakly elongate, or quite wide....................……………………………Kalcapiini
Aspidapiini Alonso-Zarazaga, 1990
Key to genera of Aspidapiini in Baltic amber
1. Scutellum acuminate at apex. Body robust............................................................. Pseudaspidapion
—Scutellum obtuse at apex. Body slender............................................................................. Baltoapion
Genus Pseudaspidapion Wanat, 1990
Type species: Apion spadiceum Wagner, 1908
Pseudaspidapion khnzoriani (Zherikhin, 1971)
Zherikhin [26]: 199, 204, Figure 2 therein, pl. X, Figure 1 therein (Apion).
Locality. Baltic amber.
Remarks. The holotype should be deposited in PIN; however, it was not found there.
Genus Baltoapion gen. nov.
urn:lsid:zoobank.org:act:C6505E9D-720B-44A2-9712-4214
Type species: Melanapion gusakovi Legalov, 2015
Diagnosis. Body length (without rostrum) 1.8–3.1 mm. Body black, slightly elongate, dorsally convex, covered with light, short hairs. Head slightly elongate. Rostrum long, slightly curved, cylindrical. Antennal scrobes weak, directed to under rostrum. Forehead flattened, wider than rostrum base. Eyes large, round, sharply convex. Temples long or short. Antennae inserted ventrally in middle or in basal one-third of rostrum. Scape elongate. Club compact, with distinct sutures between its antennomeres. Pronotum almost campaniform, roughly punctate. Scutellum slightly elongate, distinctly projecting above elytra, obtuse at apex. Elytra elongate, distinctly convex. Striae distinct and deep. Interstriae more or less wide, slightly convex, weakly finely transversely wrinkled or finely punctate. Procoxal cavities contiguous. Mesocoxal cavities separate.
Etymology. The name is formed from the Latin “balticum” (Baltic) and part of the generic name “Apion”. Gender neuter.
Comparison. The new genus differs from the Pseudaspidapion Wanat, 1990 in the scutellum obtuse at apex and more slender body.
Key to species of genus Baltoapion in Baltic amber
1. Rostrum long and thin. Antennae inserted in middle of rostrum. Temples short. Elytral interstriae finely transversely wrinkled. Body (without rostrum) smaller (1.8 mm).............. B. gusakovi
—Rostrum short and thick. Antennae inserted in basal one-third of rostrum. Temples long. Elytral interstriae finely punctate. Body (without rostrum) larger (3.1 mm)..................... B. subdiscedens
Baltoapion gusakovi (Legalov, 2015), comb. nov. (Figure 4j)
Melanapion gusakovi Legalov, 2015
Legalov [2]: 1477, 1479–1481, Figure 5a therein, plate 6, Figure 4 therein.
Locality. Baltic amber.
Remarks. This species is known only by the holotype (CVGM).
Figure 5.
Habitus of Brentidae and Curculionidae from Baltic amber. (a) Melanapion poinari, holotype, ISEA, no. BA2013/22, body, laterally. (b) M. wanati, holotype, ISEA, no. BA2012/5, body, laterally. (c) Conapium alleni, holotype, ISEA, no. BA2012/2, body, laterally. (d) Baltonanophyes crassirostre, holotype, ISEA, no. BA2017/10, body, laterally. (e,f) Dorytomus korotyaevi sp. nov., holotype, ISEA, no. BA2015/6: body, laterally, left (e); body, laterally, rigth (f). (g,h) D. bukejsi, holotype, ISEA, no. BA2019/3: body, laterally (g); body, dorsally (h). (i) D. electrinus, holotype, ISEA, no. BA2015/5, body, laterally. (j) D. nudus, holotype, ISEA, no. BA2013/5, body, laterally. (k,l) Electrotribus theryi, holotype, CCMCL, no. A6487: body, laterally (k); body, dorsally (l). Scale bars: 1.0 mm (a–j). See Section 2 for names of collections.
Figure 5.
Habitus of Brentidae and Curculionidae from Baltic amber. (a) Melanapion poinari, holotype, ISEA, no. BA2013/22, body, laterally. (b) M. wanati, holotype, ISEA, no. BA2012/5, body, laterally. (c) Conapium alleni, holotype, ISEA, no. BA2012/2, body, laterally. (d) Baltonanophyes crassirostre, holotype, ISEA, no. BA2017/10, body, laterally. (e,f) Dorytomus korotyaevi sp. nov., holotype, ISEA, no. BA2015/6: body, laterally, left (e); body, laterally, rigth (f). (g,h) D. bukejsi, holotype, ISEA, no. BA2019/3: body, laterally (g); body, dorsally (h). (i) D. electrinus, holotype, ISEA, no. BA2015/5, body, laterally. (j) D. nudus, holotype, ISEA, no. BA2013/5, body, laterally. (k,l) Electrotribus theryi, holotype, CCMCL, no. A6487: body, laterally (k); body, dorsally (l). Scale bars: 1.0 mm (a–j). See Section 2 for names of collections.

Baltoapion subdiscedens (Voss, 1953), comb. nov. (Figure 4i)
Apion subdiscedens Voss, 1953
Locality. Baltic amber.
Remarks. This species is known only by the holotype (GPIH).
Kalcapiini Alonso-Zarazaga, 1990
Key to genera of Kalcapiini in Baltic amber
1. Femora ventrally with spine at distal 1/3. Claws simple.............................................. Succinapion
—Femora without spine. Claws with teeth..........................................................................Melanapion
Genus Melanapion Wagner, 1930
Type species: Apion minimum Herbst, 1797
Subgenus Melanapionoides Legalov, 2012
Type species: Melanapion poinari Legalov, 2015
Key to species of subgenus Melanapionoides in Baltic amber
1. Eyes sharply convex. Striae of elytra relatively wide. Body relatively large, 2.4 mm long..............................................................................................................................................M. (M.) poinari
—Eyes slightly convex. Striae of elytra relatively narrow. Body relatively small, 1.7–2.0 mm long..............................................................................................................................................M. (M.) wanati
Legalov [2]: 1477–1479, Figure 4e therein, plate 6, Figure 3 therein.
Locality. Baltic amber.
Remarks. This species is known only by the holotype (ISEA).
Legalov [40]: 220, Figure 1 therein, pl. 2, Figures 3 and 4 therein.
Locality. Baltic amber.
Remarks. The holotype of this species deposited in ISEA.
Genus Succinapion Legalov et Bukejs, 2014
Type species: Succinapion telnovi Legalov et Bukejs, 2014
Succinapion telnovi Legalov et Bukejs, 2014
Legalov, Bukejs [42]: 604–606, Figures 1–3 therein.
Locality. Baltic amber.
Remarks. This species is known only by the holotype (ABCD).
Apionitae Schoenherr, 1823
Piezotrachelini Voss, 1959
Key to genera of Piezotrachelini in Baltic amber
1. Mesocoxal cavities separated. Pronotum subcylindrical................................................. Conapium
—Mesocoxal cavities contiguous. Pronotum almost campaniform............................ Baltoconapium
Genus Conapium Motschulsky, 1866
Type species: Apion gracile Gerstaecker, 1854
Subgenus Palaeoconapion Legalov, 2012
Type species: Conapium alleni Legalov, 2012
Legalov [40]: 221–222, Figure 4 therein, pl. 2, Figures 7 and 8 therein.
Locality. Baltic amber.
Remarks. This species is known only by the holotype (ISEA).
Genus Baltoconapium gen. nov.
urn:lsid:zoobank.org:act:467EEACE-C4D9-4EFE-BBBF-A16B4B8A1702
Type species: Apion anderseni Voss, 1972
Diagnosis. Body length (without rostrum) 2.0 mm. Body black, naked, lustrous. Head not constricted behind eyes. Antennae and legs brown. Rostrum weakly curved, slightly longer than pronotum. Antennal scrobes weak. Eyes large, rounded. Antennae inserted before middle. Club compact, with distinct sutures between its antennomeres. Pronotum almost campaniform, finely punctate. Elytra almost pear-shaped, stark convex, with weakly humeri. Elytral striae weak with fine points. Interstriae wide, flat, without punctures. Stria 10 absent. Prosternum not emarginate. Pro- and mesocoxal cavities contiguous. Trochanters long. Claws with teeth.
Etymology. The name is formed from the Latin “Baltic” (Baltic) and part of the generic name “Conapium”. Gender neuter.
Comparison. The new genus differs from the Conapium in the contiguous mesocoxal cavities. and almost campaniform pronotum. From the genus Piezotrachelus Schoenherr, 1839 with contiguous mesocoxal cavities it differs in the not emarginate prosternum and head not constricted behind eyes.
Baltoconapium anderseni (Voss, 1972), comb. nov. (Figure 3h)
Apion anderseni Voss, 1972
Voss [25]: 171–173, Figures 3 and 11 therein.
Locality. Baltic amber.
Remarks. This species is known only by the holotype (ZMUC).
Genus Electrapion Wagner, 1924
Type species: Apion kuntzeni Wagner, 1924
Electrapion kuntzeni (Wagner, 1924)
Wagner [27]: 134–136 (Apion).
Locality. Baltic amber.
Remarks. The genus is conditionally placed in this tribe because of the almost naked body, large size, flat scutellum, long rostrum, and elytra with nine striae. The holotype of this species is absent in ZMHB [62]. New finds of this species are needed to clarify the systematic position.
Nanophyinae Gistel, 1856
Nanophyini Gistel, 1856
Remarks. The genus Nanophyes Schoenherr, 1838 was recorded from Baltic amber [61].
Genus Baltonanophyes Legalov, 2018
Type species: Baltonanophyes crassirostre Legalov, 2018
Baltonanophyes crassirostre Legalov, 2018 (Figure 5d)
Legalov [46]: 165–166, Figure 2 therein.
Locality. Baltic amber.
Remarks. This species is known only by the holotype (ISEA).
Curculionidae Latreille, 1802
Key to subfamilies of Curculionidae in Eocene amber
1. Protibia distally with groove and row of dense erect setae.........................................Cossoninae
—Protibia distally without mouth groove and row of dense erect setae...........................................2
2. Tarsomere 5 with claws widely separated by dermal lobes. Antennae inserted at base or near middle of rostrum...................................................................................................................Dryophthorinae
—Tarsomere 5 with claws not separated by dermal lobes. Antennae inserted near middle or at rostrum apex.......................................................................................................................................................3
3. Mesepimeron enlarged and visible between bases of prosternum and elytra; if not then eyes large and subcontiguous dorsally on forehead and the maximum width of elytra usually at base..................................................................................................................................................Conoderinae
—Mesepimeron not enlarged and invisible between bases of prosternum and elytra...................4
4. Tibiae without uncus and groups of setae, sometimes with mucro. Mandibles usually massive, with scar of deciduous process....................................................................................... Entiminae
—Tibiae with uncus and two groups of setae, usually with mucro; if uncus absent, rostrum very long. Mandibles without scar.........................................................................................................................5
5. Body often with dense varnish-like coating of scales. Uncus displaced onto inner apical angle. Tibiae often with apical spurs. Ventrite 5 usually with anal setae.........................................Erirhininae
—Body without dense varnish-like coating of scales. Uncus (if present) located at outer apical angle or displaced onto inner apical angle. Tibiae without apical spurs................................................6
6. Ventrite 5 always without anal setae. Uncus absent (at least on protibiae) or not large, displaced onto inner apical angle. Mucro usually absent. Antennal scrobes usually invisible from above. Posterior angles of ventrite 2 sometimes extended strongly posteriorly toward ventrite 3.....................................................................................................................................................Curculioninae
—Ventrite 5 often with anal setae. Uncus large, located at outer apical angle or, rarely, displaced onto inner apical angle. Antennal scrobes usually visible from above. Posterior margins of ventrites 2 and 3 straight.....................................................................................................................................................7
7. Body oval or wide. Prementum without transverse groove, small. Claws free............Molytinae
—Body elongate. Prementum with transverse grooves, large. Claws usually fused at base.........................................................................................................................................................[Lixinae]
Erirhininae Schoenherr, 1825
Key to tribes of Erirhininae in Eocene amber
1. Body covered by dense varnish-like coating over scales. Antennal scrobes in anterior part seen from above. Prosternum with ventral channel......................................................................[Bagoini]
—Body naked or covered simple scales. Antennal scrobes in anterior part not seen from or visible only in place on antennal attachment. Prosternum simple..............................................................2
2. Eyes rounded. Elytral stria 9 short, fused with stria 10 at level of metacoxa. Femora with tooth..................................................................................................................................................Dorytomini
—Eyes transverse, oval, if rounded than ventrite 5 without anal setae. Elytral stria 9 long, not fused with stria 10 at level of metacoxal. Femora lacking tooth................................................[Erirhinini]
[Erirhinini Schoenherr, 1825]
Remarks. The genus Notaris Germar, 1817 was recorded from Baltic amber [61].
Dorytomini Bedel, 1886
Genus Dorytomus Germar, 1817
Type species: Curculio vorax Fabricius, 1792 (= Curculio longimanus Forster, 1771)
Key to species of genus Dorytomus in Eocene amber
1. Elytral interstriae with decumbent setae or scales............................................................................2
—Elytral interstriae without decumbent setae or scales......................................................................4
2. Elytral interstriae with decumbent setae, smooth............................................................. D. nudus
—Elytral interstriae with decumbent scales, punctate........................................................................ 3
3. Rostrum long. Body wide. Elytral interstriae slightly convex, distinctly wider than elytral striae. Pronotum finely punctate..................................................................................................... D. groehni
—Rostrum short. Body narrower. Elytral interstriae flat, equal or slightly wider than elytral striae. Pronotum coarsely punctate................................................................................. D. bukejsi sp. nov.
4. Body not lustrous, weaker convex. Pronotum and thorax densely punctate. Elytral interstriae wrinkly, punctate.............................................................................................................................. D. vlaskini
—Body lustrous, more convex. Pronotum and thorax sparsely punctate. Elytral interstriae finely punctate................................................................................................................................................... 5
5. Scape not reaching eye. Eyes coarsely faceted, with seven facets longitudinal to long axis.....................................................................................................................................................D. electrinus
—Scape reaching eye. Eyes finely faceted, with more than 15 facets longitudinal to long axis.................................................................................................................................... D. korotyaevi sp. nov.
Dorytomus bukejsi sp. nov. (Figure 5g,h)
urn:lsid:zoobank.org:act:7F6B37D3-9A6E-4E99-AB13-E90E8ED00697
Description. Male. Size. Body length (without rostrum) 2.5 mm; rostrum length 0.5 mm. Body black, covered with decumbent narrow scales. Head 0.3 times as long as rostrum. Rostrum long, subcylindrical, 5.5 times as long as wide at apex, 4.0 times as long as wide in middle and 3.6 times as long as wide at base, 1.2 times as long as pronotum, distinctly curved, densely punctate. Antennal scrobes distinct, directed to eye. Forehead 0.8 times as long as rostrum base width, flattened, punctate. Eyes large, 0.6 times as long as wide, not protruding from margin of head. Temples 0.7 times as long as eye length. Antennae inserted in apical third, geniculate. Scape 7.3 times as long as width, not reaching eye. Funicle seven-segmented. Club compact. Pronotum bell-shaped, 1.1 times as long as wide basally, 0.8 times as long as wide in middle and basally. Disc weakly convex, coarsely rugose, punctate. Elytra distinctly convex, weakly elongate, 2.9 times as long as pronotum, 1.7 times as long as wide at base, 1.5 times as long as wide in middle, 1.9 times as long as wide in apical quarter. Greatest width in middle. Humeri distinct. Striae regular, distinct and deep, with sparse deep points. Elytral interstriae weakly convex, narrow, subequal in stria width, with row of scales. Stria 9 merges with stria 10 at level of metacoxae. Thorax. Prosternum densely punctate, without postocular lobes. Metaventrite subequal to metacoxa length, weakly convex, punctate. Metepisternum narrow. Abdomen convex. Ventrites 1 and 2 quite elongate. Ventrites 3 and 4 short, equal in length. Ventrite 1 subequal to metacoxal length. Ventrite 2 1.3 times as long as ventrite 1. Ventrite 3 0.5 times as long as ventrite 2. Ventrite 5 1.7 times as long as ventrite 4, with anal setae. Legs elongate. Femora weakly clavate, with teeth. Metafemora length/width ratio 3.3. Tibiae almost straight, flattened, with small uncus displaced by inner apical angle and two apical bunches of setae. Tarsi elongate, latter with pulvilli on underside. Tarsomeres: 1 and 2—conical; 3—bilobed; 5—elongate, with large free claws with teeth. Metatarsomeres: 1—1.2 times as long as wide; 2—equal in length and width, 0.9 times as long as and equal in width to tarsomere 1; 3—equal in length and width, 1.3 times as long as and 1.3 times as wide as tarsomere 2; 5—6.5 times as long as wide, 1.6 times as long as and 0.3 times as narrow as tarsomere 3.
Material examined. Holotype (ISEA), no. BA2019/3.
Comparison. The new species differs from D. groehni in the shorter rostrum, narrower body, coarsely punctate pronotum, and elytral interstriae flat, equal or slightly wider than elytral striae.
Etymology. The epithet of this new species is dedicated to Andris Bukejs (Daugavpils) who contributed to the studies of amber Coleoptera.
Locality. Baltic amber.
Dorytomus electrinus Legalov, 2016 (Figure 5i)
Legalov [44]: 62–64, Figures 4 and 5 therein.
Locality. Baltic amber.
Remarks. This species is known only by the holotype.
Dorytomus groehni Bukejs et Legalov, 2019
Bukejs, Legalov [50], 174–177, Figures 1 and 2 therein.
Locality. Baltic amber.
Remarks. This species is known only by the holotype.
Dorytomus korotyaevi sp. nov. (Figure 5e,f)
urn:lsid:zoobank.org:act:6086E677-CFBD-4674-B0CC-BC8D9F61EE13
Description. Female. Size. Body length (without rostrum) 3.3 mm; rostrum length 0.7 mm. Body black, naked, without scales and setae. Head 0.4 times as long as rostrum. Rostrum long, subcylindrical, 4.4 times as long as wide at apex, about 4.0 times as long as wide in middle and at base, 1.3 times as long as pronotum, distinctly curved, finely punctate. Antennal scrobes distinct, directed to eye. Forehead 0.5 times as long as rostrum base width, flattened, punctate. Eyes large, not protruding from margin of head, finely faceted, with more than 15 facets longitudinal to long axis. Temples short. Antennae inserted before middle of rostrum laterally, geniculate. Scape 7.2 times as long as width, reaching eye. Funicle seven-segmented. Antennomeres: 2—1.7 times as long as wide, 0.3 times as long as and 1.2 times as wide as scape; 3—1.8 times as long as wide, 0.7 times as long as and 0.7 times as narrow as antennomere 2; 4—1.1 times as long as wide, 0.6 times as long as and 0.9 times as narrow as antennomere 3; 5—1.2 times as long as wide, 0.9 times as long as and 0.9 times as narrow as antennomere 4; 6—equal in length and width, 0.9 times as long as and 1.0 times as wide as antennomere 5; 7—0.7 times as long as wide, 1.0 times as long as and 1.5 times as wide as antennomere 6; 8—0.7 times as long as wide, 1.2 times as long as and 1.1 times as wide as antennomere 7. Club compact, 1.4 times as long as wide. Pronotum bell-shaped. Disc weakly convex, punctate. Elytra convex, weakly elongate, 4.2 times as long as pronotum. Greatest width in middle. Humeri distinct. Striae regular and distinct, with rounded and quite deep points. Elytral interstriae weakly convex, wide, 2.3–4.0 times as wide as stria width, finely punctate. Stria 9 merges with stria 10 at level of metacoxae. Thorax. Prosternum densely punctate, without postocular lobes. Precoxal part of prosternum 0.8 times as long as procoxa length. Procoxal cavities round, contiguous. Postcoxal part of prosternum 0.6 times as long as procoxa length. Mesocoxal cavities rounded, separated. Metaventrite 1.2 times as long as metacoxa length, weakly convex, punctate. Metepisternum narrow, 7.2 times as long as wide, with row of points. Abdomen convex. Ventrites oriented in one plane. Ventrites 1 and 2 quite elongate. Ventrites 3 and 4 quite short. Ventrite 1, 1.6 times as long as metacoxal length. Ventrite 2, 1.1 times as long as ventrite 1. Ventrite 3, 0.6 times as long as ventrite 2. Ventrite 4, equal in length to ventrite 3. Ventrite 5, 2.0 times as long as ventrite 4, with anal setae. Legs elongate. Femora weakly clavate, with teeth. Profemora length/width ratio 3.2. Metafemora length/width ratio 2.9. Tibiae almost straight, flattened, with small uncus displaced by inner apical angle and two apical bunches of setae. Protibiae length/width ratio 7.2. Metatibiae length/width ratio 5.0. Tarsi elongate, with pulvilli on underside. Tarsomeres: 1 and 2—conical; 3—bilobed; 5—elongate, with large free claws with teeth.
Material examined. Holotype (ISEA), BA2015/6.
Comparison. The new species differs from D. electrinus in the scape is reaching eye, and finely faceted eyes, with more than 15 facets longitudinal to long axi.
Etymology. The epithet of this new species is dedicated to Boris A. Korotyaev (Zoological Institute of the Russian Academy of Sciences, Saint-Petersburg) who contributed to the study of the genus.
Locality. Baltic amber.
Dorytomus nudus Legalov, 2016 (Figure 5j)
Legalov [45]: 978, Figure 1f therein, plate 5, Figure 3 therein.
Locality. Baltic amber.
Remarks. This species is known only by the holotype (ISEA).
Dorytomus vlaskini Legalov, Nazarenko et Perkovsky, 2019
Legalov et al. [72]: 68, 70, Figure 1b therein, plate 8, Figures 2–5 therein.
Locality. Rovno amber.
Remarks. This species is known by the holotype and two paratypes (SIZK).
Figure 6.
Habitus of Curculionidae from Baltic amber. (a–c) Electrotribus rarus sp. nov., holotype, FEH, no. 651: body, ventrally (a); body, ventro-laterally (b); prothorax and foreleg, laterally (c). (d) Anchorthorrhinus incertus, holotype, GPIH, no. 199, body, laterally. (e–g) Isalcidodes macellus, holotype, GPIH, no. 200: body, laterally, on the right (e); fore-body, laterally, on the right (f); body, laterally, on the left (g). (h) Electrotribus theryi, holotype, CCMCL, no. A6487, dorsally. (i–k) Electrorhinus friedhelmi gen. et sp. nov., holotype, FEH, no. 653, body, laterally (i); body, ventrally (k). (j) E. friedhelmi gen. et sp. nov., paratype, ISEA, no. BA2019/1, laterally. (l) Palaeodexipeus kirejtshuki, holotype, ISEA, no. BA2015/3, body, laterally. (m) Leiosoma klebsi, holotype, ISEA, no. BA2013/3, laterally. Scale bars: 1.0 mm. See Section 2 for names of collections.
Figure 6.
Habitus of Curculionidae from Baltic amber. (a–c) Electrotribus rarus sp. nov., holotype, FEH, no. 651: body, ventrally (a); body, ventro-laterally (b); prothorax and foreleg, laterally (c). (d) Anchorthorrhinus incertus, holotype, GPIH, no. 199, body, laterally. (e–g) Isalcidodes macellus, holotype, GPIH, no. 200: body, laterally, on the right (e); fore-body, laterally, on the right (f); body, laterally, on the left (g). (h) Electrotribus theryi, holotype, CCMCL, no. A6487, dorsally. (i–k) Electrorhinus friedhelmi gen. et sp. nov., holotype, FEH, no. 653, body, laterally (i); body, ventrally (k). (j) E. friedhelmi gen. et sp. nov., paratype, ISEA, no. BA2019/1, laterally. (l) Palaeodexipeus kirejtshuki, holotype, ISEA, no. BA2015/3, body, laterally. (m) Leiosoma klebsi, holotype, ISEA, no. BA2013/3, laterally. Scale bars: 1.0 mm. See Section 2 for names of collections.

[Bagoini C.G. Thomson, 1859]
Remarks. The genus Bagous Germar, 1817 was recorded from Baltic amber [58].
Molytinae Schoenherr, 1823
Key to tribes of Molytinae in Eocene amber
1. Prosternum with rostral channel bounded by carinae.....................................................................2
—Prosternum without rostral channel bounded by carinae.............................................................4
2. Postcoxal portion of prosternum bounded by carinae.................................................Aedemonini
—Postcoxal portion of prosternum not bounded by carinae..............................................................3
3. Rostral channel not reaching mesoventrite.................................................................. Sciabregmini
—Rostral channel reaching mesoventrite.................................................................. Cryptorhynchini
4. Meso- and metepisternum, meso- and metepimeron covered with dense plumose scales………………......................................................................................................................[Magdalinini]
—Meso- and metepisternum, meso- and metepimeron covered with simple scales.......................5
5. Elytral base with anterior projection near interstria 3 which extends over pronotum................................................................................................................................... [Mecysolobini]
—Elytral base straight or weakly concave.............................................................................................6
6. Metacoxae subglobular. Abdominal process broadly truncate, as wide as or wider than metacoxa. Body naked........................................................................................................................... Plintini
—Metacoxae transverse, elongate. Abdominal process acuminate, much narrower than metacoxae. Body covered with scales............................................................................................................7
7. Antennal scrobes distinctly visible from above in apical part...................................... [Molytini]
—Antennal scrobes not visible from above even in apical part..........................................................8
8. Club segment 1 with dense pubescence, not lustrous. Femora with tooth.............................................................................................................................................[Acicnemidini]
—Club segment 1 with quite sparse pubescence, lustrous. Femora without tooth........[Pissodini]
Acicnemidini Lacordaire, 1865
Genus Electrotribus Hustache, 1942
Type species: Electrotribus theryi Hustache, 1942
= Paleopissodes Ulke, 1947
Type species: Paleopissodes weigangae Ulke, 1947
= Anchorthorrhinus Voss, 1953
Type species: Anchorthorrhinus incertus Voss, 1953
= Isalcidodes Voss, 1953
Type species: Isalcidodes macellus Voss, 1953
= Protoceletes Rheinheimer, 2007
Type species: Protoceletes wolfschwenningerae Rheinheimer, 2007
Key to species of genus Electrotribus in Eocene amber
1. Procoxal cavities contiguous. Body small (2.6 mm)......................................E. wolfschwenningerae
—Procoxal cavities narrowly separated. Body large (4.1–7. mm)...................................................... 2
2. Body flattened. Scape reaching eyes........................................................................ E. rarus sp. nov.
—Body convex. Scape not reaching eyes................................................................................................3
3. Body covered with dense scales. Forehead distinctly narrower than base of rostrum................................................................................................................................................... E. theryi
—Body naked. Forehead barely narrower than base of rostrum................................. E. henningseni
Electrotribus rarus sp. nov. (Figure 6a–c)
urn:lsid:zoobank.org:act:4FB12AE0-5E4C-4FBF-AF1E-C18B3B508084
Description. Male. Size. Body length (without rostrum) 5.4 mm; rostrum length 1.2 mm. Body black, with dense appressed scales. Head. Rostrum quite long, subcylindrical, 5.0 times as long as wide at apex and base, 5.5 times as long as wide in middle, 0.9 times as long as pronotum, almost straight, punctate. Antennal scrobes directed to eye. Eyes large, not protruding from margin of head, finely faceted. Temples shorter than eye. Antennae inserted before middle of rostrum, geniculate. Scape 5.8 times as long as width, reaching eye. Antennomeres: 2—2.3 times as long as wide, 0.4 times as long as and 0.9 times as narrow as scape; 3—1.7 times as long as wide, 0.6 times as long as and 0.9 times as narrow as antennomere 2; 4–8—conical. Club compact, 2.9 times as long as wide. Pronotum bell-shaped. Disc weakly flattened, punctate. Elytra weakly flattened, elongate, 2.5 times as long as pronotum. Greatest width in middle. Humeri distinct. Striae regular and distinct. Elytral interstriae slightly wider than stria width, punctate. Stria 9 full, not merges with stria 10 at level of metacoxae. Thorax. Prosternum densely punctate, without postocular lobes. Precoxal part of prosternum 1.2 times as long as procoxa length. Procoxal cavities round, narrowly separated. Postcoxal part of prosternum 0.5 times as long as procoxa length. Mesocoxal cavities rounded, separated. Metaventrite 2.2 times as long as metacoxa length, weakly convex, punctate. Metepisternum narrow, 10.0 times as long as wide, punctate. Sclerolepidia present. Abdomen flattened. Ventrite 1, 1.7 times as long as metacoxal length. Ventrite 2, 0.9 times as long as ventrite 1. Ventrite 3, 0.3 times as long as ventrite 2. Ventrite 4, subequal in length to ventrite 3. Ventrite 5, 1.8 times as long as ventrite 4. Legs elongate. Femora weakly clavate, with teeth. Profemora length/width ratio 2.6. Metafemora length/width ratio 2.5. Apex of metafemora reaching apex of ventrite 3. Tibiae almost straight, flattened, dilated approximately in the middle, with large uncus. Protibiae length/width ratio 5.0. Metatibiae length/width ratio about 2.8. Tarsi elongate, with pulvilli on underside. Tarsomeres: 1 and 2—conical; 3—bilobed; 5—elongate, with large free claws with teeth. Protarsomeres: 1—1.1 times as long as wide; 2—0.9 times as long as wide, 0.8 times as long as and equal in width to tarsomere 1; 3—0.8 times as long as wide, 1.5 times as long as and 1.6 times as wide as tarsomere 2; 5—2.8 times as long as wide, 1.2 times as long as and 0.4 times as narrow as tarsomere 3.
Material examined. Holotype (FEH), no. 651.
Comparison. The new species differs from E. theryi in the flattened body and scape reaching eye.
Etymology. The name of this new species is formed from the Latin word “rarus” (rare).
Locality. Baltic amber.
Electrotribus henningseni (Voss, 1972), comb. nov.
Pissodes henningseni Voss, 1972
Voss [25]: 180, Figures 10 and 14 therein.
Locality. Baltic amber.
Remarks. Holotype was not found in the ZMUC. This species should belong to the genus Electrotribus due to the narrowly separated procoxal cavities, tooth on the femora, and densely punctured pronotum. The structure of the claws is not known. This species differs from E. theryi in the wider forehead and body without scales.
Hustache [28]: 109, Figure 1 therein.
= Paleopissodes weigangae Ulke [29]: 2–4, Figures 1–4 therein, syn. nov.
= Anchorthorrhinus incertus Voss [24]: 132–133, Figures 9a and 10a therein.
= Isalcidodes macellus Voss [24]: 134–135, Figures 9b and 10b therein.
Locality. Baltic amber.
Remarks. Holotypes are kept in CCMCL (Electrotribus theryi Hustache, 1942), ANSP (Paleopissodes weigangae Ulke, 1947), and GPIH (Anchorthorrhinus incertus Voss, 1953 and Isalcidodes macellus Voss, 1953). Kuschel [90] established synonymy for Paleopissodes weigangae, Anchorthorrhinus incertus (Figure 6d) and Isalcidodes macellus (Figure 6e–g), but he did not study the type of Electrotribus theryi (Figure 5k–l and Figure 6h). Study of photographs, and comparative material showed that Paleopissodes weigangae syn. nov. is synonymous with Electrotribus theryi. Thanks to the help of Dr. A. Nel (Paris), I managed to obtain photographs of the type of Electrotribus theryi from the CCMCL and taken by Didier Berthet (Lyon).
Electrotribus wolfschwenningerae (Rheinheimer, 2007)
Rheinheimer [34]: 16–18, Figure 13 therein (Protoceletes).
Locality. Baltic amber.
Remarks. This species is known only by the holotype (SMNS).
[Pissodini Gistel, 1856]
Remarks. The genus Pissodes Germar, 1817 was recorded from Baltic amber [57].
[Magdalinini Pascoe, 1870]
Remarks. The genus Magdalis Germar, 1817 was recorded from Baltic amber [61].
[Molytini Schoenherr, 1823]
Remarks. The genus Hylobius Germar, 1817 was recorded from Baltic amber [72].
Plinthini Lacordaire, 1863
Leiosomatina Reitter, 1913
Genus Leiosoma Stephens, 1829
Type species: Curculio punctatus Marsham, 1802 non Scopoli, 1763 (= Curculio deflexus Panzer, 1795)
Leiosoma klebsi Legalov, 2016 (Figure 6m)
Legalov [45]: 970, 974, Figure 1a therein, Plate 4, Figure 1 therein.
Locality. Baltic amber.
Remarks. This species is known only by the holotype (ISEA).
[Mecysolobini Reitter, 1913]
Remarks. The genus Alcidodes Marshall, 1939 was recorded from Baltic amber [62].
Sciabregmini Legalov, Kirejtshuk et Nel, 2019
Genus Sciabregma Scudder, 1893
Type species: Sciabregma rugosum Scudder, 1893
Sciabregma squamosa Legalov, Kirejtshuk et Nel, 2019
Legalov et al. [20]: 70–71, Figure 1c,g therein, plate 18, Figures 2–4 therein.
Locality. Oise amber.
Remarks. This species is known only by the holotype (MNHN).
Aedemonini Faust, 1898
Remarks. It is the first record of the tribe in Baltic amber.
Genus Electrorhinus gen. nov.
urn:lsid:zoobank.org:act:F2947E0E-0534-4C6C-A226-3F40689E4FFF
Type species: Electrorhinus friedhelmi sp. nov.
Diagnosis. Body covered with dense decumbent scales. Rostrum quite long, subequal in length to pronotum. Eyes weakly protruding from margin of head, coarsely faceted. Antennae inserted near middle of rostrum, geniculate. Club quite long. Pronotum coarsely rugose, punctate. Elytra weakly convex, with distinct humeri. Elytral stria 9 full. Prosternum with weak postocular lobes. Prosternum with rostral channel bounded by carinae. Apex of rostral channel bounded by carina. Rostral channel reaching mesoventrite. Postcoxal portion of prosternum bounded by carinae. Metaventrite equal in length to metacoxa length, about five times longer than ventrite 3. Sclerolepidia present. Ventrites 1 and 2 long. Ventrites 3 and 4 short. Femora weakly clavate, sulcate beneath, with teeth. Metafemora reaching ventrite 3. Tarsal claws simple.
Comparison. The new genus differs from the recent genus Rhadinomerus Faust, 1892 by the femora sulcate beneath, shorter metafemora reaching only ventrite 3 and short ventrites 3 and 4.
Etymology. The name of the new genus is formed from the Latin “electrum” (amber) and the Greek “rhinos” (nose). Gender masculine.
Remarks. The new genus belongs to Aedemonini, as it has the prosternum with a rostral channel bounded by the carinae, apex of the rostral channel bounded by the carina, the rostral channel reaching the mesoventrite, postcoxal portion of the prosternum bounded by the carinae and present sclerolepidia.
Electrorhinus friedhelmi sp. nov. (Figure 6i–k)
urn:lsid:zoobank.org:act:2D5A7817-2721-4E56-9848-F02417CDDBED
Description. Male. Size. Body length (without rostrum) 5.7–6.2 mm; rostrum length 1.3–1.4 mm. Body black, with dense decumbent quite narrow scales. Head. Rostrum quite long, 3.8 times as long as wide at apex, 4.2 times as long as wide in middle, 3.5 times as long as wide at base, 0.9–1.1 times as long as pronotum, weakly curved, punctate. Antennal scrobes directed under rostrum. Eyes large, weakly protruding from margin of head, coarsely faceted. Temples short. Antennae inserted near middle of rostrum, geniculate. Scape 7.0 times as long as width, not reaching eye. Funicle with long conical segments. Antennomeres: 2—4.9 times as long as wide, 0.5 times as long as and 0.7 times as narrow as scape; 3—4.0 times as long as wide, 0.6 times as long as and 0.7 times as narrow as antennomere 2. Club compact, 3.0 times as long as wide. Pronotum bell-shaped. Disc weakly convex, coarsely rugose-punctate. Elytra weakly convex, quite elongate, 2.8–2.9 times as long as pronotum. Greatest width in middle. Humeri distinct. Striae regular and distinct. Elytral interstriae wide, punctate, 2.3–3.2 times as long as width of striae. Stria 9 full, not merges with stria 10 at level of metacoxae. Thorax. Prosternum densely punctate, with weak postocular lobes. Precoxal part of prosternum 1.3 times as long as procoxa length. Procoxal cavities round, separated. Postcoxal part of prosternum 0.7 times as long as procoxa length. Mesocoxal cavities rounded, separated. Metaventrite convex, equal in length to metacoxa length, 4.9 times as long as ventrite 3, weakly convex, punctate. Metepisternum narrow, 10.8 times as long as wide, punctate. Sclerolepidia present. Abdomen flattened. Ventrite 1, slightly wider than metacoxal length. Ventrite 2, 1.3 times as long as ventrite 1. Ventrite 3, 0.2 times as long as ventrite 2. Ventrite 4, equal in length to ventrite 3. Ventrite 5, 4.3 times as long as ventrite 4. Legs elongate. Femora weakly clavate, sulcate beneath, with teeth. Profemora length/width ratio 3.6. Mesofemora length/width ratio 3.1–3.5. Metafemora length/width ratio 3.7, reaching ventrite 3. Tibiae weakly curved, flattened, with uncus. Protibiae length/width ratio 7.1. Metatibiae length/width ratio 4.8–5.2. Tarsi elongate, with pulvilli on underside. Tarsomeres: 1 and 2—conical; 3—bilobed; 5—elongate, with large free claws without teeth. Metatarsomeres: 1—2.5 times as long as wide; 2—1.4 times as long as wide, 0.6 times as long as and equal in width to tarsomere 1; 3—0.7 times as long as wide, equal in length as and 2.0 times as wide as tarsomere 2; 5—5.7 times as long as wide, 1.5 times as long as and 0.2 times as wide as tarsomere 3.
Material examined. Holotype (FEH), no. 653 and paratype (ISEA), no. BA2019/1.
Etymology. The epithet of this new species is dedicated to Friedhelm Eichmann (Hannover) who provided one specimen of this species for study.
Locality. Baltic amber.
Cryptorhynchini Schoenherr, 1826
Key to subtribes of Cryptorhynchini in Eocene amber
1. Metaventrite short, usually shorter than ventrite 3. Abdominal process as broad as coxa. Metepisternum narrow or partly hidden, or metepisternal sutures absent. Elytra with more or less reduced humeri.................................................................................................................................[Tylodina]
—Metaventrite more or less elongate, longer than ventrite 3. Abdominal process narrower than coxa. Metepisternum distinct, broader. Elytra often with rectangular humeri....................................................................................................................................Cryptorhynchina
Cryptorhynchina Schoenherr, 1826
Remarks. The genus Cryptorhynchus Illiger, 1807 was recorded from Baltic amber [61].
Key to genera of Cryptorhynchina in Eocene amber
1. All femora without teeth........................................................................................................Oisecalles
—All femora with teeth.........................................................................................................Succinacalles
Genus Oisecalles Legalov, Kirejtshuk et Nel, 2019
Type species: Oisecalles latosquamosus Legalov, Kirejtshuk et Nel, 2019
Oisecalles latosquamosus Legalov, Kirejtshuk et Nel, 2019
Legalov et al. [20]: 72, Figure 2a,b,d,e therein, plate 18, Figures 5 and 6 therein; plate 18, Figure 1 therein.
Locality. Oise amber.
Remarks. This species is known only by the holotype (MNHN).
Genus Succinacalles Zherikhin, 1971
Type species: Succinacalles uniqus Zherikhin, 1971
Succinacalles uniqus Zherikhin, 1971
Zherikhin [26]: 199, 207–208, Figure 5 therein, pl. X., Figure 3a,b therein.
Locality. Baltic amber.
Remarks. The holotype should be deposited in PIN; however, it was not found there.
[Tylodina Lacordaire, 1865]
Remarks. The genus Acalles Schoenherr, 1825 was recorded from Baltic amber [61].
[Lixinae Schoenherr, 1823]
[Lixini Schoenherr, 1823]
Remarks. The genus Lixus Fabricius, 1801 was recorded from Baltic amber [61].
Dryophthorinae Schoenherr, 1825
Key tribes of Dryophthorinae in Eocene amber
1. Procoxal cavities contiguous. Tarsi with very small fourth tarsomere...................... Stromboscerini
—Procoxal cavities separated. Tarsi with distinct five tarsomeres............................ Dryophthorini
Dryophthorini Schoenherr, 1825
Remarks. The genus Dryophthorus Germar, 1824 was recorded from Baltic amber [61].
Genus Rhinoporkus Legalov, Kirejtshuk et Nel, 2019
Type species: Rhinoporkus gratiosus Legalov, Kirejtshuk et Nel, 2019
Rhinoporkus gratiosus Legalov, Kirejtshuk et Nel, 2019
Legalov et al. [20]: 74, 76, Figure 2c,f,g therein, plate 19, Figures 2–5 therein.
Locality. Oise amber.
Remarks. This species is known only by the holotype (MNHN).
Stromboscerini Lacordaire, 1865
Key to genera of Stromboscerini in Eocene amber
1. Funicle four-segmented..................................................................................................... Rovnoslonik
—Funicle six-segmented.........................................................................................................................2
2. Elytral interstriae carinate..............................................................................................Palaeodexipeus
—Elytral interstriae convex.....................................................................................Stenommatomorphus
Genus Palaeodexipeus Legalov, 2016
Type species: Palaeodexipeus kirejtshuki Legalov, 2016
Palaeodexipeus kirejtshuki Legalov, 2016 (Figure 6l)
Legalov [44]: 60–62, Figures 2 and 3 therein.
Locality. Baltic amber.
Remarks. This species is known only by the holotype (ISEA).
Genus Rovnoslonik Legalov, Nazarenko et Perkovsky, 2019
Type species: Rovnoslonik damzeni Legalov, Nazarenko et Perkovsky, 2019
Rovnoslonik damzeni Legalov, Nazarenko et Perkovsky, 2019
Legalov et al. [72]: 64, 66, Figure 1a,g therein, pl. 7, Figures 1–4 therein.
Locality. Rovno amber.
Remarks. This species is known only by the holotype (ISEA).
Genus Stenommatomorphus Nazarenko, 2009
Type species: Stenommatomorphus hexarthrus Nazarenko, 2009
Stenommatomorphus hexarthrus Nazarenko, 2009
Nazarenko, Perkovsky [66]: 1100, Figure 1 therein, pl. III, Figures 1–3 therein.
Locality. Rovno amber.
Remarks. This species is known only by the holotype (SIZK).
Cossoninae Schoenherr, 1825
Key to tribes of Cossoninae in Eocene amber
1. Mesoventrite more or less strongly depressed below level of metaventrite.............[Rhyncolini]
—Mesoventrite on level of metaventrite................................................................................................2
2. Procoxal cavities narrowly separated. Humeri rounded..............................................Dryotribini
—Procoxal cavities widely separated. Humeri distinct.....................................................[Cossonini]
[Rhyncolini Gistel, 1856]
Remarks. The genera Rhyncolus Germar, 1817 and Choerorhinus Fairmaire, 1858 were recorded from Baltic amber [61].
Dryotribini LeConte, 1876
Key to genera of Dryotribini in Eocene amber
1. Funicle 4–6-segmented.........................................................................................................................2
—Funicle seven-segmented.....................................................................................................................3
2. Funicle four-segmented...........................................................................................Necrodryophthorus
—Funicle five-segmented..................................................................................................Synommatodes
—Funicle six-segmented..................................................................................................Electrocossonus
3. Rostrum thick and straight, 1.8 times as long as wide in middle........................Ampharthropelma
—Rostrum slender and curved, 2.7–6.9 times as long as wide in middle........................Caulophilus
Genus Necrodryophthorus Voss, 1953
Type species: Necrodryophthorus inquilinus Voss, 1953
Necrodryophthorus inquilinus Voss, 1953
Voss [24]: 137, Figure 11 therein.
Locality. Baltic amber.
Remarks. This species is known only by the holotype (GPIH).
Genus Synommatodes Voss, 1953
Type species: Synommatus patruelis Voss, 1953
Synommatodes patruelis (Voss, 1953) (Figure 7f)
Voss [24]: 138, Figure 12 therein (Synommatus).
Locality. Baltic amber.
Remarks. The holotype of this species deposited in GPIH.
Genus Electrocossonus gen. nov.
urn:lsid:zoobank.org:act:144A9B84-F5C9-4FD0-BA65-65AE67C55E61
Type species: Electrocossonus kirejtshuki sp. nov.
Diagnosis. Body covered with decumbent narrow scales. Rostrum shorter than pronotum, without secondary scrobes. Antennal scrobes directed under rostrum. Eyes coarsely faceted, only slightly protruding from margin of head. Antennae inserted in middle of rostrum. Antennomere quite long, not reaching eye. Funicle six-segmented. Antennomere 2 long, conical. Antennomeres 3–8 wide, conical. Antennal club compact. Elytra quite robust. Humeri weakly smoothed. Elytral interstriae as wide as or narrower than width of striae. Prothorax with distinct postocular lobes. Procoxal cavities narrowly separated. Suture between metepisternum and metaventrite strongly convex. Abdomen flattened. Ventrites 1, 2, and 5, long. Ventrites 3 and 4, short. Femora clavate, without tooth. Tarsi elongate, with pulvilli on underside. Tarsomere 3 weakly bilobed. Claws free, simple.
Etymology. The name is formed from the Latin “electrum” (amber) and generic name “Cossonus”. Gender masculine.
Comparison. The new genus differs from other genera of the tribe in the six-segmented funicle. From the genus Caulophilus, it also differs in a more robust body and strongly convex suture between the metepisternum and metaventrite.
Remarks. The new genus belongs to Cossoninae, as it has the protibiae with a notch in the distal half and a row of dense erect setae, metatibiae lacking an apical group of setae and tarsomere 3 weakly bilobate. The rostrum is longer that its width and narrowly separated cavities indicate affinity to Dryotribini.
Electrocossonus kirejtshuki sp. nov. (Figure 7b,c)
urn:lsid:zoobank.org:act:E65A36B5-B3CD-4F1A-B377-D08ECB3A4E66
Description. Male. Size. Body length (without rostrum) 2.4 mm. Rostrum length 0.5 mm. Body black, covered with decumbent narrow scales. Head densely punctate. Rostrum quite long, about 0.7 times as long as pronotum, 4.5 times as long as width at apex, 3.0 times as long as width at mid-rostrum and at base, distinctly curved, punctate. Mandibles small. Antennal scrobes deep, lateral, directed under rostrum. Secondary scrobes absent. Forehead quite wide. Eyes rounded, large, coarsely faceted, almost not protruding from margin of head. Vertex weakly convex, densely punctate. Temples short. Antennae inserted in middle of rostrum, laterally. Antennomeres: 1—long, 2.6 times as long as wide, not reaching eye; 2—long-conical, 2.2 times as long as wide, 0.6 times as long as and about 0.7 times as narrow as scape; 3–8—wide, conical; 3—0.7 times as long as wide, 0.2 times as long as and 0.7 times as narrow as antennomere 2; 4—0.8 times as long as wide, 1.3 times as long as and 1.1 times as wide as antennomere 3; 5—0.7 times as long as wide, 0.8 times as long as and 0.9 times as narrow as antennomere 4; 6—0.6 times as long as wide, 0.9 times as long as and 1.1 times as wide as antennomere 5; 7—0.7 times as long as wide, 1.4 times as long as and 1.2 times as wide as antennomere 6. Antennal club compact, about 1.8 times as long as wide. Pronotum almost bell-shaped. Disc flattened, densely punctate. Intervals between points narrower than diameter of points. Elytra quite robust, convex, 1.6 times as long as pronotum. Humeri weakly smoothed. Striae deep, distinct, regular and wide. Stria 9 not shortened. Interstriae quite narrow, weakly convex, 0.6–1.0 times as long as width of striae. Thorax. Prothorax with distinct postocular lobes, coarsely punctate. Precoxal part of prosternum 1.1 times as long as procoxa length. Procoxal cavities rounded, narrowly separated. Postcoxal part of prosternum 0.7 times as long as procoxa length. Mesocoxal cavities rounded and separated. Metaventrite 1.9 times as long as length of mesocoxa, flattened, densely punctate. Suture between metepisternum and metaventrite strongly convex. Metepisternum 3.1 times as long as wide in middle. Metacoxal cavities dilated, separated by apex of ventrite 1. Abdomen flattened. Ventrite 1, 1.3 times as long as metacoxae. Ventrite 2, 0.9 times as long as ventrite 1. Ventrite 3, 0.5 times as long as ventrite 2. Ventrite 4, equal in length to ventrite 3. Ventrite 5, 2.5 times as long as ventrite 4. Legs quite long. Procoxae spherical. Femora clavate, without tooth. Profemora 3.0 times as long as wide in middle. Metafemora 3.6 times as long as wide in middle. Tibiae almost straight, weakly flattened, with large uncus. Protibia 4.0 times as long as wide in middle, with notch in distal half, and row of dense erect setae. Metatibia 4.1 times as long as wide in middle, lacking apical bunch of setae. Tarsi elongate, with pulvilli on underside. Tarsomeres: 1 and 2—conical; 3—weakly bilobed; 5—elongate. Claws large, free, without teeth.
Material examined. Holotype (ISEA), no. BA2017/5.
Etymology. The epithet of this new species is dedicated to entomologist Dr. Alexander G. Kirejtshuk (Zoological Institute of the Russian Academy of Sciences, Saint-Petersburg).
Locality. Baltic amber.
Genus Ampharthropelma Voss, 1972
Type species: Ampharthropelma decipiens Voss, 1972
Ampharthropelma decipiens Voss, 1972 (Figure 7a,h)
Voss [25]: 177–178, Figures 7, 8 and 13 therein.
Locality. Baltic amber.
Remarks. This species is known only by the holotype (ZMUC).
Genus Caulophilus Wollaston, 1854
Type species: Caulophilus sculpturatus Wollaston, 1854 (= Rhyncolus oryzae Gyllenhal, 1838)
Key to species of genus Caulophilus in Eocene amber
1. Body covered with setaceous scales……………………………………............................……..…. 2
—Body covered with narrow scales………………………………………............................…………3
2. Rostrum slender, distinctly curved. Pronotum distinctly narrower than base of elytra……………………........................................................................................................................ C. rarus
—Rostrum robust, weakly curved. Pronotum slightly narrower than base of elytra .……….......……................................................................................................................... C. sucinopunctatus
3. Body (without rostrum) smaller, body length 1.6 mm. Elytra 2.0 times as long as wide medially ……..................................................................................................................................C. squamosus
—Body (without rostrum) larger, body length 1.9–2.3 mm. Elytra 2.2–2.3 times as long as wide medially...............................................................................................................................................................4
4. Punctures in elytral striae round. Elytra with fine hardly noticeable setae. Humeri more strongly smoothed. Body (without rostrum) smaller, body length 1.9 mm....................... C. martynovae
—Punctures in elytral striae oval or rounded rectangular. Elytra with distinct setae. Humeri weakly smoothed. Body (without rostrum) larger, body length 2.2 mm...............……....... C. zherikhini
Caulophilus martynovae Legalov, Nazarenko et Perkovsky, 2019
Legalov et al. [72]: 66, 68, Figure 1c,d therein, plate 7, Figure 5 therein; plate 8, Figure 1 therein.
Locality. Rovno amber.
Remarks. This species is known only by the holotype (SIZK).
Caulophilus rarus Legalov, 2016 (Figure 7e)
Legalov [45]: 974–975, Figure 1b therein, plate 4, Figure 2 therein.
Locality. Baltic amber.
Remarks. This species is known only by the holotype (ISEA).
Caulophilus squamosus Legalov, 2016 (Figure 7d)
Legalov [45]: 975, Figure 1c therein, plate 4, Figure 3 therein.
Locality. Baltic amber.
Remarks. This species is known only by the holotype (ISEA).
Caulophilus sucinopunctatus (Kuska, 1992) (Figure 7g,i)
Kuska [31]: 112, Figures 6–9 therein (Phloeophagus).
Locality. Baltic amber.
Remarks. Holotype is kept in the EIW.
Figure 7.
Habitus of Curculionidae from Baltic amber. (a,h) Ampharthropelma decipiens, holotype, ZMUC, no. 958: body, laterally (a); body, dorsally (h). (b,c) Electrocossonus kirejtshuki, holotype, ISEA, no. BA2017/5: body, laterally, on the left (b); body, laterally, on the right (c). (d) Caulophilus squamosus, holotype, ISEA, no. BA2012/9, body, laterally. (e) C. rarus, holotype, ISEA, no. BA2011/5, body, laterally. (f) Synommatodes patruelis, holotype, GPIH, no. 202, body, laterally. (g) Caulophilus sucinopunctatus, holotype, EIW, no. 554, body, dorsally. (i) Caulophilus sucinopunctatus, specimen, ISEA, no. BA2011/3, body, laterally. (j) Ceutorhynchus succinus, holotype, ISEA, no. BA2012/3, body, laterally. (k) C. electrinus, holotype, ISEA, no. BA2012/10, body, laterally. (l) C. alekseevi, holotype, CVIA, no AWI-005, body, dorsally. Scale bars: 1.0 mm. See Section 2 for names of collections.
Figure 7.
Habitus of Curculionidae from Baltic amber. (a,h) Ampharthropelma decipiens, holotype, ZMUC, no. 958: body, laterally (a); body, dorsally (h). (b,c) Electrocossonus kirejtshuki, holotype, ISEA, no. BA2017/5: body, laterally, on the left (b); body, laterally, on the right (c). (d) Caulophilus squamosus, holotype, ISEA, no. BA2012/9, body, laterally. (e) C. rarus, holotype, ISEA, no. BA2011/5, body, laterally. (f) Synommatodes patruelis, holotype, GPIH, no. 202, body, laterally. (g) Caulophilus sucinopunctatus, holotype, EIW, no. 554, body, dorsally. (i) Caulophilus sucinopunctatus, specimen, ISEA, no. BA2011/3, body, laterally. (j) Ceutorhynchus succinus, holotype, ISEA, no. BA2012/3, body, laterally. (k) C. electrinus, holotype, ISEA, no. BA2012/10, body, laterally. (l) C. alekseevi, holotype, CVIA, no AWI-005, body, dorsally. Scale bars: 1.0 mm. See Section 2 for names of collections.

Caulophilus zherikhini Nazarenko, Legalov et Perkovsky, 2011
Nazarenko et al. [67]: 288, 290, Figure 1 therein, pl. 7, Figures 1–9 therein.
Locality. Rovno amber.
Remarks. This species is known only by the holotype (SIZK).
[Cossonini Schoenherr, 1825]
Remarks. The genus Mesites Schoenherr, 1838 was recorded from Baltic amber [61].
Conoderinae Schoenherr, 1833
Key to supertribes of Conoderinae in Eocene amber
1. Eyes large, subcontiguous dorsally separated by very narrow forehead..........................................................................................................................................Conoderitae
—Eyes not large. Forehead more or less wide. Body more or less rounded, snout.........................................................................................................................................Ceutorhynchitae
Ceutorhynchitae Gistel, 1848
Key to tribes of Ceutorhynchitae in Eocene amber
1. Rostrum quite short, no more or slightly longer than 3× wide at apex, wider than width of profemur or equal in width to.........................................................................................................Phytobiini
—Rostrum distinctly longer than 3× wide at apex, narrower than width of profemur....................................................................................................................................Ceutorhynchini
[Phytobiini Gistel, 1856]
Remarks. The genus Rhinoncus Schoenherr, 1825 was recorded from Baltic amber [61].
Ceutorhynchini Gistel, 1848
Key to genera of Ceutorhynchini in Baltic amber
1. Meso- and metaventrite without rostral channel. Body piceous............................. Ceutorhynchus
—Meso- and metaventrite with rostral channel. Body ferruginous............................. Baltocoeliodes
Genus Ceutorhynchus Germar, 1824
Type species: Curculio assimilis Paykull, 1792
Key to species of genus Ceutorhynchus in Baltic amber
1. Tarsomere 5 long. Ventrite 5 with long erect hairs apically. Pronotum with deep medial groove.................................................................................................................................................C. alekseevi
—Tarsomere 5 rather short. Ventrite 5 without long erect hairs apically. Pronotum with weak longitudinal medial groove..............................................................................................................................2
2. Intervals of elytra with one or two rows of tubercles. Punctures of pronotum larger, with shorter scales. Femora with large teeth........................................................................................C. electrinus
—Intervals of elytra punctate. Punctures of pronotum smaller, with longer scales. Intervals narrower, with longer and more erect scales. Femora with small teeth...................................C. succinus
Ceutorhynchus alekseevi Legalov, 2016 (Figure 7l)
Legalov [45]: 976–977, Figure 1e therein, plate 4, Figure 5 therein.
Locality. Baltic amber.
Remarks. This species is known by the holotype (SIZK).
Ceutorhynchus electrinus Legalov, 2016 (Figure 7k,j)
Legalov [45]: 976, Figure 1d therein, plate 4, Figure 4 therein.
Locality. Baltic amber.
Remarks. This species is known by the holotype (ISEA).
Ceutorhynchus succinus Legalov, 2013 (Figure 7k)
Legalov [41]: 73–74, Figures 25 and 26 therein.
Locality. Baltic amber.
Remarks. This species is known by the holotype (ISEA).
Genus Baltocoeliodes Legalov et Bukejs, 2018
Type species: Baltocoeliodes sontagae Legalov et Bukejs, 2018
Baltocoeliodes sontagae Legalov et Bukejs, 2018
Legalov, Bukejs [47]: 186–189, Figures 1 and 2 therein.
Locality. Baltic amber.
Remarks. This species is known only by the holotype (MAIG).
Conoderitae Schoenherr, 1833
Remarks. The specimen of “tropischen Zygopinae” was recorded from Baltic amber [62].
Conoderini Schoenherr, 1833
Genus Jantarhinus Legalov, Kirejtshuk et Nel, 2019
Type species: Jantarhinus compressus Legalov, Kirejtshuk et Nel, 2019
Jantarhinus compressus Legalov, Kirejtshuk et Nel, 2019
Legalov et al. [20]: 76–77, Figures 2h and 3a,b therein, plate 19, Figure 6 therein; Figures 1 and 2 therein.
Locality. Oise amber.
Remarks. This species is known only by the holotype (MNHN).
Curculioninae Latreille, 1802
Key to tribes of Curculioninae in Eocene amber
1. Funicle five-segmented........................................................................................................ [Mecinini]
—Funicle seven- or six-segmented.........................................................................................................2
2. Metatibiae narrowed apically, with oblique apical comb of setae. Eyes large, strongly prominent or confluent on forehead............................................................................................. Rhamphini
—Metatibiae not narrowed apically, with transverse apical comb of setae. Eyes simple...............3
3. Mandibles vertically articulated.....................................................................................Curculionini
—Mandibles horizontal articulated........................................................................................................4
4. Posterior margins of ventrites 2–4 curved distinctly posteriad on lateral sides. Antennal scrobes directed under eyes................................................................................................................Tychiini
—Posterior margins of ventrites 2–4 straight. Antennal scrobes directed to eyes...........................5
5. Precoxal portion of prosternum short..................................................................... [Anthonomini]
—Precoxal portion of prosternum elongate...........................................................................................6
6. Pygidium covered by elytra. Tibiae uncinate...................................................................... Ellescini
—Pygidium exposed. Tibiae without uncus..........................................................................................7
7. Antennal scrobes oblique directed toward the rostrum base. Antennae inserted close to rostrum apex. Eyes very large........................................................................................................Eugnomini
—Antennal scrobes directed to eyes. Antennae inserted close to rostrum middle. Eyes small....................................................................................................................................................Acalyptini
Acalyptini Thompson, 1859
Genus Jantaronosik Legalov, Kirejtshuk et Nel, 2019
Type species: Jantaronosik nebulosus Legalov, Kirejtshuk et Nel, 2019
Jantaronosik nebulosus Legalov, Kirejtshuk et Nel, 2019
Legalov et al. [20]: 78, Figure 3d,e therein, plate 20, Figures 3 and 4 therein.
Locality. Oise amber.
Remarks. This species is known only by the holotype (MNHN).
Curculionini Latreille, 1802
Remarks. The key to species is based on the key from Pelsue and O’Brien [91].
Key to subtribes of Curculionini in Baltic amber
1. Tarsal claws connate, simple. Trochanter with stiff bristle...............................................Erganiina
—Tarsal claws free, appendiculate or not. Trochanter without stiff bristle......................................2
2. Tarsal claws simple. Pygidium covered by elytra in males...............................................Timolina
—Tarsal claws appendiculate. Pygidium exposed in males.......................................[Curculionina]
Erganiina Pelsue et O’Brien, 2011
Genus Pseudoergania Legalov, 2019
Type species: Pseudoergania perkovskyi Legalov, 2019
Pseudoergania perkovskyi Legalov, 2019 (Figure 8c)
Legalov [51]: 56–57, pl. 5 therein.
Locality. Baltic amber.
Remarks. This species is known only by the holotype (ISEA).
Timolina Heller, 1925
Genus Baltocurculio gen. nov.
urn:lsid:zoobank.org:act:E8B220C6-6553-4AD0-AA30-B14E49ECC18A
Type species: Baltocurculio manukyani sp. nov.
Diagnosis. Body large, covered with appressed and decumbent scales. Rostrum cylindrical, very long. Mandibles articulated in vertical plane, triangular. Antennal scrobes lateral, directed to eye. Eyes oval, quite small. Antennae inserted in middle of rostrum. Scape not reaching eye. Funicle seven-segmented. Antennomere 3 longer than antennomere 2. Antennal club compact. Pronotum almost bell-shaped. Base of pronotum biconcave. Scutellum convex, longer than wide. Elytra almost trapezoidal. Lateral margin of elytra not sinuate above metepisternum. Humeri distinctly convex. Greatest width in humeri. Stria 9 not shortened. Interstriae wide and convex. Prothorax without postocular lobes. Pre- and postcoxal parts of prosternum short. Metepisternum quite narrow. Abdomen weakly flattened. Ventrites 1 and 2 not elongate, and ventrites 3 and 4 not short. Pygidium covered by elytra. Femora clavate, with large tooth. Metafemora slightly enlarged, reaching apex of abdomen. Pro- and mesotibiae with mucro. Claws weakly diverging, free, without teeth.
Etymology. The name is formed from the Latin “balticum” (Baltic) and generic name “Curculio”. Gender neuter.
Comparison. The new genus differs from the African genus Timola Pascoe, 1886 in the metafemora reaching apex of abdomen, small eyes, not shortened ventrite 3 and 4, elongate body and long rostrum. It is distinguished from the South American genus Megaoculis Pelsue et O’Brien, 2011 by the quite simple claws, small eyes, very long rostrum, slightly enlarged metafemur and body without erect scales.
Remarks. Mandibles are vertically articulated, rostrum long and thin, and femora with teeth suggest that the new genus belongs to Curculionini. Simple free tarsal claws and pygidium covered by elytra in male support its assignment to Timolina.
Baltocurculio manukyani sp. nov. (Figure 8a,b)
urn:lsid:zoobank.org:act:C67306A3-968A-4EEC-8A80-14CB8F18E5A7
Description. Male. Size. Body length (without rostrum) 6.6 mm. Rostrum length 3.1 mm. Body black, covered with dense appressed and sparse decumbent brownish scales. Head almost spherical, transversely wrinkled ventrally. Rostrum almost cylindrical, long, about 1.7 times as long as pronotum, about 16.0 times as long as width at apex and at base, about 13.3 times as long as wide at mid-rostrum, distinctly curved, finely punctate. Mandibles articulated in vertical plane, small, triangular. Antennal scrobes lateral, directed to eye. Forehead quite wide, about 1.6 times as wide as rostrum base width, convex, densely punctate. Eyes oval, quite small, weakly convex, not protruding from margin of head. Vertex weakly convex, densely punctate. Temples quite short. Antennae inserted in middle of rostrum, laterally. Antennomeres: 1—long, about 7.0 times as long as wide, not reaching eye; 2–8 (funicle)—conical; 2—about 1.8 times as long as wide, about 0.2 times as long as and about 0.8 times as narrow as scape; 3—about 2.9 times as long as wide, about 1.4 times as long as and about 0.9 times as narrow as antennomere 2; 4—about 1.8 times as long as wide, about 0.6 times as long as and about 0.9 times as narrow as antennomere 3; 5—about 2.0 times as long as wide, subequal in length and about 0.9 times as narrow as antennomere 4; 6—about 2.0 times as long as wide, about 0.7 times as long as and about 0.7 times as narrow as antennomere 5; 7—about 1.6 times as long as wide, subequal in length and about 1.3 times as wide as antennomere 6; 8—about 0.8 times as long as wide, about 0.8 times as long as and about 1.6 times as wide as antennomere 7. Antennal club compact, about 1.6 times as long as wide. Pronotum almost bell-shaped, about 1.4 times as long as wide apically, little longer than wide in middle and basally. Sides weakly rounded in first half and almost subparallel in second half. Disc flattened, densely punctate. Base of pronotum biconcave. Mesonotum. Scutellum convex, about 1.8 times as long as wide. Elytra almost trapezoidal, weakly convex, about 2.7 times as long as pronotum, about 1.7 times as long as wide basally, about 2.4 times as long as wide in middle, about 3.5 times as long as wide in apical quarter. Lateral margin of elytra not sinuate above metepisternum. Humeri convex. Greatest width in humeri. Striae deep, distinct, regular and narrow. Stria 9 not shortened, not merged with stria 10 at level of metacoxae. Interstriae quite wide, weakly convex, 2.5–4.0 times as long as width of striae. Elytral apices rounded separately. Thorax. Prothorax without postocular lobes, coarsely punctate. Pre- and postcoxal parts of prosternum short, subequal in length. Precoxal part of prosternum about 0.4 times as long as procoxa length. Procoxal cavities rounded, contiguous. Mesocoxal cavities rounded and separated. Metaventrite slightly longer than length of mesocoxa, weakly convex, densely punctate. Metepisternum about 9.6 times as long as wide in middle. Metacoxal cavities dilated, subequal in length to metaventrite, separated by apex of ventrite 1. Abdomen weakly flattened. Ventrite 1 about 0.4 times as long as metacoxae. Ventrite 2 about 1.6 times as long as ventrite 1. Ventrite 3 about 1.2 times as long as ventrite 2. Ventrite 4 about 0.8 times as long as ventrite 3. Ventrite 5 about 1.4 times as long as ventrite 4. Pygidium covered by elytra. Legs elongate. Procoxae conical. Femora clavate, with large tooth in apical third. Profemora about 4.4 times as long as wide after tooth. Metafemora about 6.0 times as long as wide after tooth. Tibiae almost straight, weakly flattened, weakly widened apically. Pro- and mesotibiae with mucro. Protibia about 7.6 times as long as wide in middle. Metatibia about 7.4 times as long as wide in middle. Tarsi elongate, with pulvilli on underside. Tarsomeres: 1 and 2—conical; 3—bilobed; 5—elongate. Claws large, diverging, free, without teeth. Protarsomeres: 1—about 1.6 times as long as wide; 2—about 1.1 times as long as wide, about 0.7 times as long as and about 1.1 times as wide as tarsomere 1; 3—subequal in length and width, about 1.6 times as long as and about 1.7 times as wide as tarsomere 2; 5—about 5.7 times as long as wide, about 1.3 times as long as and about 0.2 times as narrow as tarsomere 3.
Material examined. Holotype (KRAM), no. BX 100-19.
Etymology. The epithet of this new species is dedicated to paleoentomologist Dr. Andranik R. Manukyan (Kaliningrad Regional Amber Museum).
Locality. Baltic amber.
[Curculionina Latreille, 1802]
Remarks. The genus Balaninus Germar, 1817 (=Curculio Linnaeus, 1758) was recorded from Baltic amber [61]. Representatives of this subtribe were not found by the author, but the probability of finding them is high, because eight species of oak are described from Baltic amber [92].
Ellescini C.G. Thomson, 1859
Key to genera of Ellescini in Baltic amber
1. Stria 9 not shortened. Body covered with appressed and decumbent scales.....Succinostyphlus
—Stria 9 short, fused with stria 10 at level of metacoxae. Body covered with appressed scales................................................................................................................................................Pachytychius
Genus Succinostyphlus Kuska, 1996
Type species: Succinostyphlus mroczkowskii Kuska, 1996
Remarks. Probably the record of the genus Pseudostyphlus Tournier, 1874 from Baltic amber [61] concerns to this genus.
Succinostyphlus mroczkowskii Kuska, 1996 (Figure 8d,l,m)
Kuska [32]: 15, Figure 8 therein.
= Electrotribus erectosquamata Rheinheimer, 2007, syn. nov.
Figure 8.
Habitus of Curculioninae from Baltic amber. (a,b) Baltocurculio manukyani gen. et sp. nov., holotype, KRAM, no. BX 100-19: body, dorsally (a); body, ventrally (b). (c) Pseudoergania perkovskyi, holotype, ISEA, no. BA2017/7, body, laterally. (d) Succinostyphlus mroczkowskii, holotype, EIW, no. 6427, body, dorsally. (e) Archaeoeugnomus balticus, holotype, ISEA, no. BA2012/15, body, laterally. (f) Anthonoeugnomus barsevskisi gen. et sp. nov., holotype, ISEA, no. BA2017/2, body, laterally. (g,h) Pachytychius eocenicus, holotype, ISEA, no. BA2011/7: body, dorso-laterally (g); body, laterally (h). (i) Mazurieugnomus pilosus gen. et sp. nov., holotype, ISEA, no. 2016/1, body, laterally. (j,k) Groehnius parvum sp. nov., holotype, FEH, no. 649: body, dorso-laterally (j); body, dorsally (k). (l) Succinostyphlus mroczkowskii, specimen, ISEA, no. 2012/20, body laterally. (m) S. mroczkowskii, GPIH, no. SEBS00292a, body, laterally. Scale bars: 1.0 mm. See Section 2 for names of collections.
Figure 8.
Habitus of Curculioninae from Baltic amber. (a,b) Baltocurculio manukyani gen. et sp. nov., holotype, KRAM, no. BX 100-19: body, dorsally (a); body, ventrally (b). (c) Pseudoergania perkovskyi, holotype, ISEA, no. BA2017/7, body, laterally. (d) Succinostyphlus mroczkowskii, holotype, EIW, no. 6427, body, dorsally. (e) Archaeoeugnomus balticus, holotype, ISEA, no. BA2012/15, body, laterally. (f) Anthonoeugnomus barsevskisi gen. et sp. nov., holotype, ISEA, no. BA2017/2, body, laterally. (g,h) Pachytychius eocenicus, holotype, ISEA, no. BA2011/7: body, dorso-laterally (g); body, laterally (h). (i) Mazurieugnomus pilosus gen. et sp. nov., holotype, ISEA, no. 2016/1, body, laterally. (j,k) Groehnius parvum sp. nov., holotype, FEH, no. 649: body, dorso-laterally (j); body, dorsally (k). (l) Succinostyphlus mroczkowskii, specimen, ISEA, no. 2012/20, body laterally. (m) S. mroczkowskii, GPIH, no. SEBS00292a, body, laterally. Scale bars: 1.0 mm. See Section 2 for names of collections.

Locality. Baltic amber.
Remarks. Holotype of Succinostyphlus mroczkowskii Kuska, 1996 is kept in the EIW and holotype of Electrotribus erectosquamata Rheinheimer, 2007 in SMNS. Study of descriptions, images, and comparative material showed that Electrotribus erectosquamata syn. nov. is synonymous with Succinostyphlus mroczkowskii.
Genus Pachytychius Jekel, 1861
Type species: Pachytychius eocenicus Legalov, 2016
Pachytychius eocenicus Legalov, 2016 (Figure 8g,h)
Legalov [45]: 980, Figure 1g therein, plate 5, Figure 4 therein.
Locality. Baltic amber.
Remarks. This species is known only by the holotype (ISEA).
[Anthonomini C. G. Thomson, 1859]
Remarks. The genus Anthonomus Germar, 1817 was recorded from Baltic amber [61].
Eugnomini Lacordaire, 1863
Key to genera of Eugnomini in Baltic amber
1. Femora without ventral teeth. Scape not reaching the anterior margin of eye..............Groehnius
—Femora with ventral teeth. Scape reaching the anterior margin of eye..........................................2
2. Apices of eyes are located in same plane with forehead. Antennae inserted beyond middle.................................................................................................................. Anthonoeugnomus gen. nov.
—Eyes protruding above forehead. Antennae inserted in apical one-third of rostrum..................3
3. Rostrum equal to head and pronotum combined. Tooth on metafemora large. Antennal club simple, oval. Intervals interstriae weakly convex..............................................................Archaeoeugnomus
—Rostrum longer than head and pronotum combined. Tooth on metafemora small. Antennal club attenuate. Intervals interstriae distinctly convex................................... Mazurieugnomus gen. nov.
Genus Archaeoeugnomus Legalov, 2016
Type species: Archaeoeugnomus balticus Legalov, 2016
Archaeoeugnomus balticus Legalov, 2016 (Figure 8e)
Legalov [45]: 981, Figure 1h therein, plate 5, Figure 5 therein.
Locality. Baltic amber.
Remarks. This species is known only by the holotype (ISEA).
Genus Anthonoeugnomus gen. nov.
urn:lsid:zoobank.org:act:47C24C02-B76A-48D6-BAA4-73E093A85E7A
Type species: Anthonoeugnomus barsevskisisp. nov.
Diagnosis. Body covered with subdecumbent hairs. Rostrum long, curved, longer than head and pronotum combined. Antennal scrobes oblique, directed toward rostrum base. Eyes convex, large. Apices of eyes are located in same plane with forehead. Temples short. Antennae inserted beyond middle of rostrum. Scape reaching eye. Club compact. Pronotum coarsely rugose, punctate. Elytra elongate. Interstriae convex. Precoxal part of prosternum elongate. Ventrites 1 and 2 quite long. Ventrites 3 and 4 quite short. Metafemora with large teeth. Claws free, simple.
Etymology. The name is formed from part of the generic name “Anthomonus” and generic name “Eugnomus”. Gender masculine.
Comparison. The new genus differs from the other genera from Baltic amber in the apices of the eyes are located in same plane with the forehead and antennae inserted beyond middle of the rostrum. It differs from Mazurieugnomus in the large tooth on the metafemora, weakly convex elytral interstriae, a large pronotum and elytra without long decumbent setae.
Anthonoeugnomus barsevskisi sp. nov. (Figure 8f)
urn:lsid:zoobank.org:act:6E3FAFA2-3E77-4759-BAB4-A3155C04C19D
Description. Female. Size. Body length (without rostrum) 4.1 mm; rostrum length 1.1 mm. Body black, covered with quite short pale sparse subdecumbent hairs. Head. Rostrum long, distinctly curved, 2.5 times as long as pronotum, longer than head and pronotum combined, 8.3 times as long as wide apically and medially, 6.3 times as long as wide basally, densely punctate. Antennal scrobes oblique, lateral, directed toward rostrum base. Forehead quite narrow, flattened. Eyes distinctly convex and oval, very large. Temples 0.5 times as long as eye. Antennae inserted beyond middle of rostrum. Scape elongate, reaching eye, 11.8 times as long as wide. Antennomeres: 2–8—long, conical; 2—2.3 times as long as wide, 0.2 times as long as and 0.9 times as narrow as scape; 3—2.5 times as long as wide, 0.6 times as long as and 0.6 times as narrow as antennomere 2: 4–6—subequal; 4—2.0 times as long as wide, 0.8 times as long as and equal in width to antennomere 3; 7—1.4 times as long as wide, 0.9 times as long as and 1.3 times as wide as antennomere 6; 8—1.5 times as long as wide, 1.3 times as long as and 1.2 times as wide as antennomere 7. Club compact, 1.7 times as long as wide, 0.6 times as long as funicle. Pronotum coarsely rugose, punctate, flattened. Elytra elongate, 4.2 times as long as pronotum. Humeri weakly convex. Striae wide and deep. Stria 9 short, fused with stria 10 at level of metacoxae. Interstriae convex, 1.7–2.8 times as wide as striae. Thorax. Prothorax punctate. Precoxal part of prosternum elongate, 1.6 times as long as procoxal length. Postcoxal part of prosternum 0.4 times as long as procoxal length. Metaventrite weakly convex, densely punctate, 1.9 times as long as metacoxal length. Metepisternum quite narrow, 3.1 times as long as wide medially, punctate. Abdomen convex, punctate. Ventrite 1, 1.4 times as long as metacoxae. Ventrite 2, 1.2 times as long as ventrite 1. Ventrites 3 and 4, equal in length. Ventrite 3, 0.4 times as long as ventrite 2. Ventrite 5, 1.7 times as long as ventrite 4. Legs elongate. Procoxae almost conical. Mesocoxae rounded, separated. Metacoxae elongate, shorter than ventrite 1. Femora swollen, punctate, with large teeth. Profemora 3.2 times as long as wide before tooth. Metafemora 3.1 times as long as wide before tooth. Tibiae elongate, almost straight, without mucro and uncus. Metatibiae 8.6 times as long as wide in middle. Tarsi elongate. Tarsomeres: 1—elongate; 2—conical; 3—bilobed; 5—elongate. Claws free, without teeth. Protarsomeres: 1—1.6 times as long as wide; 2—equal in length and width, 0.7 times as long as and 1.1 times as wide as tarsomere 1; 3—0.8 times as long as wide, equal in length and 1.3 times as wide as tarsomere 2; 5—3.3 times as long as wide, 1.3 times as long as and 0.3 times as narrow as tarsomere 3. Metatarsomeres: 1—2.4 times as long as wide; 2—1.3 times as long as wide, 0.7 times as long as and 1.2 times as wide as tarsomere 1; 3—0.7 times as long as wide, equal in length and 1.8 times as wide as tarsomere 2.
Material examined. Holotype (ISEA), BA2017/2.
Etymology. The epithet of this new species is dedicated to Arvids Barsevskis (Daugavpils) who made a great contribution to the organization of work on the studies of beetles in the Baltic region.
Locality. Baltic amber.
Genus Mazurieugnomus gen. nov.
urn:lsid:zoobank.org:act:2FC1E7B2-C2E0-49C9-9266-0022C4A0683F
Type species: Mazurieugnomus pilosussp. nov.
Diagnosis. Body covered with long decumbent hairs. Rostrum long, curved, longer than head and pronotum combined. Eyes protruding above forehead. Antennal scrobes oblique, directed toward rostrum base. Forehead quite narrow. Temples quite long. Antennae inserted in apical one-third of rostrum. Club compact, attenuate. Pronotum coarsely punctate. Elytral interstriae distinctly convex. Precoxal part of prosternum long. Ventrites 1 and 3 quite long. Ventrites 3 and 4 shorter. Femora with small teeth. Claws free, without teeth.
Etymology. The epithet of this new species is deicated to Miłosz A. Mazur (Opole University) and generic name “Eugnomus”. Gender masculine.
Comparison. The new genus differs from the Archaeoeugnomus in the rostrum longer than head and pronotum combined, small tooth on the metafemora, attenuate antennal club, and distinctly convex intervals interstriae.
Mazurieugnomus pilosus sp. nov. (Figure 8i)
urn:lsid:zoobank.org:act:CEE44BB1-2596-40FD-960D-DC681CB74AB8
Description. Male. Size. Body length (without rostrum) 2.5 mm; rostrum length 1.4 mm. Body black, covered with long decumbent hairs. Head. Rostrum long, weakly curved, 1.9 times as long as pronotum, longer than head and pronotum combined, 9.4 times as long as wide apically, 6.6 times as long as wide medially, 6.0 times as long as wide basally, punctate. Antennal scrobes oblique, directed toward rostrum base. Forehead quite narrow. Eyes distinctly convex and oval, very large. Temples quite long. Antennae inserted in apical one-third of rostrum. Scape elongate, reaching eye, 15.8 times as long as wide. Antennomeres: 2–8—long, conical; 2—2.7 times as long as wide; 3—2.5 times as long as wide, 0.5 times as long as and 0.5 times as narrow as antennomere 2; 4—1.8 times as long as wide, 0.7 times as long as and equal in width to antennomere 3; 5—1.7 times as long as wide, 0.9 times as long as and 0.9 times as narrow as antennomere 4; 6—1.8 times as long as wide, 0.9 times as long as and 0.8 times as narrow as antennomere 5; 7—subequal to sixth; 8—0.8 times as long as wide, 0.6 times as long as and 1.6 times as wide as antennomere 7. Club compact, 3.8 times as long as wide. Pronotum coarsely punctate, flattened. Elytra elongate, 3.9 times as long as pronotum. Humeri weakly convex. Striae wide and deep. Stria 9 short, fused with stria 10 at level of metacoxae. Interstriae convex, about 1.7 times as wide as striae. Thorax. Prothorax densely punctate. Precoxal part of prosternum 1.5 times as long as procoxal length. Postcoxal part of prosternum 0.4 times as long as procoxal length. Metaventrite convex, densely punctate, 1.7 times as long as metacoxal length. Metepisternum quite narrow. Abdomen weakly convex, punctate. Ventrite 1, about 2.3 times as long as metacoxae. Ventrite 2, 0.8 times as long as ventrite 1. Ventrite 3, 0.6 times as long as ventrite 2. Ventrite 4, 0.9 times as long as ventrite 3. Ventrite 5, 1.8 times as long as ventrite 4. Legs elongate. Procoxae almost conical. Mesocoxae rounded, separated. Femora swollen, punctate, with teeth. Profemora about 6.5 times as long as wide before tooth. Mesofemora about 5.8 times as long as wide before tooth. Metafemora about 5.1 times as long as wide before tooth, with small tooth. Tibiae elongate, almost straight, without mucro and uncus. Protibiae about 6.1 times as long as wide in middle. Mesotibiae about 8.0 times as long as wide in middle. Metatibiae about 7.3 times as long as wide in middle. Tarsi elongate. Tarsomeres: 1—elongate; 2—conical; 3—bilobed; 5—elongate. Claws free, without teeth. Mesotarsomeres: 1—2.1 times as long as wide; 2—1.3 times as long as wide, 0.8 times as long as and 1.2 times as wide as tarsomere 1; 3—0.7 times as long as wide, 0.8 times as long as and 1.6 times as wide as tarsomere 2; 5—4.7 times as long as wide, 1.4 times as long as and 0.2 times as narrow as tarsomere 3.
Material examined. Holotype (ISEA), no. 2016/1.
Etymology. The epithet of this new species is formed from the Latin “pilosus” (hairy).
Locality. Baltic amber.
Genus Groehnius Bukejs et Legalov, 2019
Type species: Groehnius electrum Bukejs et Legalov, 2019
Key to species of genus Groehnius in Baltic amber
1. Large (3.5 mm). Pronotum distinctly narrowed before apex. Rostrum without middle carina. Elytral interstriae much wider than striae................................................................................... G. electrum
—Small (1.7 mm). Pronotum slightly narrowed before apex. Rostrum with middle carina. Elytral interstriae slightly wider or equal to the width of striae................................... G. parvum sp. nov.
Groehnius electrum Bukejs et Legalov, 2019
Bukejs, Legalov [49]: 46–48, Figures 1 and 2 therein.
Locality. Baltic amber.
Remarks. This species is known only by the holotype (GPIHG).
Groehnius parvum sp. nov. (Figure 8j,k)
urn:lsid:zoobank.org:act:7440EE5C-786D-4CAD-9465-A738AABE32FE
Description. Male. Size. Body length (without rostrum) 1.7 mm; rostrum length 0.4 mm. Body black, covered with long decumbent hairs. Head. Rostrum long, weakly curved, slightly longer than pronotum, 3.3 times as long as wide apically, 3.8 times as long as wide medially, 2.9 times as long as wide basally, finely punctate, middle carina. Antennal scrobes oblique, directed toward rostrum base. Forehead slightly narrower than rostrum base. Eyes distinctly convex, very large. Temples 0.5 times as long as eye length. Antennae inserted in apical one-fourth of rostrum. Scape elongate, not reaching eye, 9.5 times as long as wide. Antennomeres: 2–8—conical; 2—2.3 times as long as wide, 0.2 times as long as and 0.8 times as narrow as scape; 3—2.5 times as long as wide, 0.7 times as long as and 0.7 times as narrow as antennomere 2; 4—1.5 times as long as wide, 0.6 times as long as and equal in width to antennomere 3; 5—1.2 times as long as wide, equal in length and 1.3 times as wide as antennomere 4; 6—equal in length and width, 0.8 times as long as and equal in width to antennomere 5; 7—1.5 times as long as wide, 1.8 times as long as and 1.2 times as wide as antennomere 6; 8—1.3 times as long as wide, 0.9 times as long as and equal in width to antennomere 7. Club compact, 3.2 times as long as wide, 0.7 times as long as funicle. Pronotum slightly narrowed before apex, 0.9 as long as wide apically, 0.8 times as long as wide medially and basally, coarsely punctate, flattened. Mesonotum. Scutellum small, almost triangular, longer than wide. Elytra elongate, 4.0 times as long as pronotum, 1.8 times as long as wide basally, 1.9 times as long as wide in middle, 2.5 times as long as wide in apical quarter. Humeri weakly convex. Striae wide and deep. Interstriae convex, narrow, 1.0–1.5 times as wide as striae. Thorax. Prothorax punctate. Metaventrite weakly convex, punctate. Metepisternum quite narrow. Abdomen weakly convex, punctate. Ventrites 1 and 2 long. Legs elongate. Procoxae almost conical. Mesocoxae rounded, separated. Femora swollen, punctate, without teeth. Tibiae elongate, almost straight, without mucro and uncus. Tarsi elongate. Tarsomeres: 1—elongate; 2—conical; 3—bilobed; 5—elongate. Claws free, without teeth.
Material examined. Holotype (FEH), no. 649.
Comparison. The new species differs from G. electrum in the smaller body size, pronotum slightly narrowed before apex, rostrum width, middle carina and elytral interstriae slightly wider or equal to the width of striae.
Etymology. The epithet of this new species is formed from the Latin “parvum” (small).
Locality. Baltic amber.
Rhamphini Rafinesque, 1815
Key to subtribe of Rhamphini in Baltic amber
1. Metafemora nondilated. All femora with large tooth near middle. Scape elongate. Posterior margins of ventrites 2–4 curved weakly posteriad on lateral sides............................. Palaeorhamphina
—Metafemora dilated. All femora without teeth or femora with small teeth. Scape short. Posterior margins of ventrites 2–4 curved distinctly posteriad on lateral sides................................................................................................................................................. Rhamphina
Palaeorhamphina Legalov, 216
Genus Palaeorhamphus Legalov, 2016
Type species: Palaeorhamphus primitivus Legalov, 2016
Key to species of genus Palaeorhamphus in Baltic amber
1. Forehead narrow. Rostrum slender.................................................................................P. primitivus
—Forehead quite wide. Rostrum more robust......................................................................................2
2. Pronotal punctation finer and sparse. Base of elytra slightly concave....... P. eichmanni sp. nov.
—Pronotal punctation larger and dense. Base of elytra strongly concave......... P. damzeni sp. nov.
Palaeorhamphus damzeni sp. nov. (Figure 9c,d)
urn:lsid:zoobank.org:act:1CA22E90-E780-4789-8FE3-2784B077B889
Description. Female. Size. Body length (without rostrum) 2.3 mm. Rostrum length 0.5 mm. Body black, covered with long, pale, erect setae. Head. Rostrum cylindrical, weakly curved, without carinae, shorter than pronotum, 3.1 times as long as wide apically, 2.7 times as long as wide medially, 3.1 times as long as wide basally, finely punctate. Antennal scrobes lateral and directed toward rostrum base. Forehead flat, 0.6 times as narrow as rostrum basally, punctate. Eyes rounded, not protruding from margin of head. Vertex weakly convex, densely punctate. Temples distinctly shorter than eye. Antennae. Scape elongate, reaching eye. Antennal club compact. Pronotum bell-shaped, 1.3 times as long as wide apically, slightly wider than length in middle, 0.9 times as long as wide basally, densely and quite coarsely punctate. Interspaces between punctures slightly smaller than puncture diameter. Sides of pronotum weakly convex in middle. Mesonotum. Scutellum 1.3 times as long as wide. Elytra wide, convex, 2.0 times as long as pronotum, 1.3 times as long as wide basally, 0.9 times as long as wide in middle, 1.9 times as long as wide in apical quarter. Base of elytra strongly concave. Humeri absent. Elytral striae deep, distinct, and regular, with quite rounded, and dense punctures. Interstriae with 1–3 rows of erect setae, 2.7–3.0 times as wide as striae. Thorax. Prothorax coarsely punctate, without postocular lobes. Precoxal part of prosternum 0.3 times as long as procoxae. Metepisternum 4.3 times as long as wide, densely punctate. Abdomen densely punctate. Ventrite 1, 1.8 times as long as metacoxae. Ventrite 2, 0.9 times as long as ventrite 1. Posterior margins of ventrites 2–4 curved posteriad on lateral sides. Ventrites 3 and 4, equal in length. Ventrite 3, 0.4 times as long as ventrite 2. Ventrite 5, 2.2 times as long as ventrite 4. Legs elongate. Femora punctate, with tooth. Tibiae weakly curved, weakly flattened, and dilated toward apex. Tarsomeres: 1—conical; 2—widely conical; 3—widely bilobed; 5—elongate. Claws large, diverging, with teeth. Mesotarsomeres: 1—1.2 times as long as wide; 2—0.9 times as long as wide, 0.9 times as long as and 1.2 times as wide as tarsomere 1; 3—0.7 times as long as wide, 1.2 times as long as and 1.6 times as wide as tarsomere 2; 5—3.7 times as long as wide, 1.4 times as long as and 0.3 times as narrow as tarsomere 3.
Material examined. Holotype (ISEA), no. BA2019/4.
Comparison. The new species differs from P. eichmanni sp. nov. in the large, dense pronotal punctation and strongly concave elytral base.
Etymology. The epithet of this new species is dedicated to Jonas Damzen (Vilnius) who sold this specimen.
Locality. Baltic amber.
Palaeorhamphus eichmanni sp. nov. (Figure 9a,b)
urn:lsid:zoobank.org:act:0B9A0D8A-B13A-4666-911C-44B946D8E669
Description. Female. Size. Body length (without rostrum) 2.5 mm. Rostrum length 0.5 mm. Body brown, covered with long, erect setae. Head. Rostrum subcylindrical, weakly curved, without carinae, shorter than pronotum, finely punctate. Antennal scrobes lateral and directed toward rostrum base. Forehead flat, narrower than rostrum base, densely punctate. Eyes rounded, weakly protruding from margin of head. Vertex weakly convex, densely punctate. Temples shorter than eye. Antennae. Scape elongate, reaching eye. Antennal club compact and elongate. Pronotum bell-shaped, 1.2 times as long as wide apically, 0.9 times as long as wide in middle, 0.7 times as long as wide basally, quite finely and sparsely punctate. Interspaces between punctures longer than puncture diameter. Sides of pronotum widened to base. Mesonotum. Scutellum 0.8 times as long as wide. Elytra wide, convex, 2.6 times as long as pronotum, 1.3 times as long as wide basally and in middle, 2.2 times as long as wide in apical quarter. Base of elytra weakly concave. Humeri smoothed. Elytral striae deep, distinct, and regular. Interstriae with row of erect setae, 2.0–4.0 times as wide as striae. Thorax. Prothorax coarsely punctate, without postocular lobes. Precoxal part of prosternum 0.4 times as long as procoxae. Metaventrite 1.2 times as long as metacoxal length. Metepisternum 9.0 times as long as wide, densely punctate. Abdomen punctate. Ventrite 1, 1.2 times as long as metacoxae. Ventrite 2, equal to ventrite 1. Posterior margins of ventrites 2–4 curved posteriad on lateral sides. Ventrites 3 and 4, subequal in length. Ventrite 3, 0.5 times as long as ventrite 2. Ventrite 5, 1.5 times as long as ventrite 4. Legs elongate. Femora punctate, with tooth. Tibiae weakly curved, weakly flattened, and dilated toward apex. Tarsomeres: 1—conical; 2—widely conical; 3—widely bilobed; 5—elongate. Claws large, diverging, with teeth.
Material examined. Holotype (FEH), no. 652.
Comparison. The new species differs from P. damzeni sp. nov. in the finer and sparse pronotal punctation and slightlyconcave elytral base. It differs from P. primitivus in the quite wide forehead and more robust rostrum.
Figure 9.
Habitus of Palaeorhamphus spp. from Baltic amber. (a,b) P. eichmanni sp. nov., holotype, FEH, no. 652: body, dorsally (a); body, ventrally (b). (c,d) P. damzeni sp. nov., holotype, ISEA, no. BA2019/4: body, dorsally (c); body, ventrally (d). (e,f) P. primitivus, holotype, ISEA, no. BA2012/12: body, ventrally (e); body, ventro-laterally (f). Scale bars: 1.0 mm. See Section 2 for names of collections.
Figure 9.
Habitus of Palaeorhamphus spp. from Baltic amber. (a,b) P. eichmanni sp. nov., holotype, FEH, no. 652: body, dorsally (a); body, ventrally (b). (c,d) P. damzeni sp. nov., holotype, ISEA, no. BA2019/4: body, dorsally (c); body, ventrally (d). (e,f) P. primitivus, holotype, ISEA, no. BA2012/12: body, ventrally (e); body, ventro-laterally (f). Scale bars: 1.0 mm. See Section 2 for names of collections.

Etymology. The epithet of this new species is dedicated to Friedhelm Eichmann (Hannover) who provided this specimen for study.
Locality. Baltic amber.
Palaeorhamphus primitivus Legalov, 2016 (Figure 9e,f)
Legalov [45]: 982–983, Figure 2a therein, plate 5, Figure 6 therein.
Locality. Baltic amber.
Remarks. This species is known only by the holotype (ISEA) and paratype (CAGB).
Rhamphina Rafinesque, 1815
Key to genera of Rhamphina in Baltic amber
1. Flagellum six-segmented. Pronotum with erect setae.....................................................Tachyerges
—Flagellum seven-segmented. Pronotum without erect setae............................................ Orchestes
Genus Orchestes Illiger, 1798
Type species: Orchestes signifer Creutzer, 1799 (= Curculio avellanae Donovan, 1797)
Orchestes tatjanae Legalov, 2016 (Figure 10a,b)
Legalov [45]: 983–984, Figure 2b therein, plate 5, Figure 7 therein.
Locality. Baltic amber.
Remarks. This species is known only by the holotype (ISEA).
Genus Tachyerges Schoenherr, 1825
Type species: Curculio solicis Linnaeus, 1758
Tachyerges hyperoche Legalov et Poinar, in lit.
Legalov, Poinar [53], in litteris.
Locality. Baltic amber.
Remarks. This species is known only by the holotype (ISEA).
[Mecinini Gistel, 1856]
Remarks. The genus Mecinus Germar, 1821 was recorded from Baltic amber [58].
Tychiini C.G. Thomson, 1859
Genus Eocenesibinia Legalov, 2015
Type species: Eocenesibinia prussica Legalov, 2016
Eocenesibinia prussica Legalov, 2016 (Figure 10c)
Legalov [45]: 984–985, Figure 2c therein, plate 5, Figure 8 therein.
Locality. Baltic amber.
Remarks. This species is known only by the holotype (ISEA).
Camarotini Schoenherr, 1833
Remarks. It is the first record of Camarotini in Baltic amber.
Figure 10.
Habitus of Curculionidae from Baltic amber. (a,b) Orchestes tatjanae, holotype, ISEA, no. BA2014/4: body, laterally, on the left (a); body, laterally, on the right (b). (c) Eocenesibinia prussica, holotype, ISEA, no. BA2013/4, body, laterally. (d,g) Paleodontopus smirnovae gen. et sp. nov., holotype, KRAM, no. 6504: body, ventro-laterally (d); body, laterally (g). (e,f) Palaeophelypera kuscheli, holotype, ISEA, no. BA2012/4: body, dorsally (e); body, laterally (f). (h,i) Limalophus poinari sp. nov., holotype, ISEA, no. BA2017/6: rostrum and head, laterally (h); body, laterally (i). Scale bars: 1.0 mm. See Section 2 for names of collections.
Figure 10.
Habitus of Curculionidae from Baltic amber. (a,b) Orchestes tatjanae, holotype, ISEA, no. BA2014/4: body, laterally, on the left (a); body, laterally, on the right (b). (c) Eocenesibinia prussica, holotype, ISEA, no. BA2013/4, body, laterally. (d,g) Paleodontopus smirnovae gen. et sp. nov., holotype, KRAM, no. 6504: body, ventro-laterally (d); body, laterally (g). (e,f) Palaeophelypera kuscheli, holotype, ISEA, no. BA2012/4: body, dorsally (e); body, laterally (f). (h,i) Limalophus poinari sp. nov., holotype, ISEA, no. BA2017/6: rostrum and head, laterally (h); body, laterally (i). Scale bars: 1.0 mm. See Section 2 for names of collections.

Prionomerina Lacordaire, 1863
Genus Paleodontopus gen. nov.
urn:lsid:zoobank.org:act:8C0A461D-8433-4B9C-8ED4-4A3786E1A0CF
Type species: Paleodontopus smirnovae sp. nov.
Diagnosis. Body convex, covered with curved setae. Rostrum subcylindrical, distinctly longer than pronotum. Antennal scrobes lateral. Forehead narrow. Eyes not large and not protruding from margin of head. Scape reaching eye. Pronotum and elytra quite wide. Elytral stria 9 full. Pre- and postcoxal parts of prosternum short. Procoxal cavities contiguous. Posterior margins of ventrites 2–4 curved posteriad on lateral sides. Profemora not enlarged, with large serrate tooth. Meso- and metafemora with simple tooth. Claws large, diverging, with teeth.
Comparison. The new genus similar to the genus Odontopus Say, 1831 but differs in the dentate metafemora, eyes not protruding from a margin of the head, and not enlarged profemora.
Etymology. The name of this new genus is formed from the Greek “palaios” (ancient) and generic name Odontopus. Gender masculine.
Remarks. The new genus belongs to Camarotini because its profemora contains a large serrate tooth. Contiguous procoxal cavities and not flattened body indicate its attribution to Prionomerina.
Paleodontopus smirnovae sp. nov. (Figure 10d,g)
urn:lsid:zoobank.org:act:553BBDA0-7673-48E4-8D6F-67D23E8C7461
Description. Female. Size. Body length (without rostrum) 4.0 mm. Rostrum length 1.3 mm. Body black, lustrous, covered with long, curved setae. Head. Rostrum subcylindrical, weakly curved, without carinae, 1.7 times as long as pronotum, 5.3 times as long as wide apically, 4.9 times as long as wide medially, finely punctate. Antennal scrobes lateral and directed toward rostrum base. Forehead flat, 0.3 times as narrow as rostrum base, punctate. Eyes rounded, not protruding from margin of head. Vertex weakly convex, finely punctate. Temples distinctly shorter than eye. Antennae. Scape elongate, reaching eye. Antennal club compact, 2.3 times as long as wide. Pronotum bell-shaped, 0.9 times as long as wide apically, 0.7 times as long as wide in middle and basally, coarsely punctate. Interspaces between punctures slightly smaller than puncture diameter. Sides of pronotum weakly convex. Elytra wide, convex, 3.4 times as long as pronotum, 1.2 times as long as wide basally, 1.1 times as long as wide in middle. Base of elytra almost straight. Humeri distinct. Elytral striae deep, distinct, and regular. Stria 9 full, not merges with striae 10 near metacoxa. Interstriae with 1–2 rows of curved setae, 2.0–2.7 times as wide as striae. Thorax. Prothorax densely punctate, without postocular lobes. Pre- and postcoxal parts of prosternum short. Metaventrite weakly convex, densely punctate, 1.3 times as long as metacoxal length. Metepisternum 6.0 times as long as wide, coarsely punctate. Abdomen punctate, weakly flattened. Ventrite 1, 0.8 times as long as metacoxae. Ventrite 2, equal in length to ventrite 1. Posterior margins of ventrites 2–4 curved posteriad on lateral sides. Ventrites 3 and 4, equal in length. Ventrite 3, 0.8 times as long as ventrite 2. Ventrite 5, 1.5 times as long as ventrite 4. Legs elongate. Femora clavate. Profemora not enlarged, with large serrate tooth, 3.8 times as long as wide before tooth. Meso- and metafemora with simple tooth. Mesofemora 3.8 times as long as wide before tooth. Metafemora 3.5 times as long as wide before tooth. Tibiae weakly curved, weakly flattened, with uncus. Tarsomeres: 1 and 2—conical; 3—widely bilobed; 5—elongate. Claws large, diverging, with teeth. Protarsomeres: 1—2.6 times as long as wide; 2—equal in length and width, 0.5 times as long as and 1.2 times as wide as tarsomere 1; 3—equal in length and width, 1.7 times as long as and 1.7 times as wide as tarsomere 2; 5—3.0 times as long as wide, 0.9 times as long as and 0.3 times as narrow as tarsomere 3.
Material examined. Holotype (KRAM), no. 6504.
Etymology. The epithet of this new species is dedicated to Anna V. Smirnova (Kaliningrad Regional Amber Museum), who helped the author in the studies of Baltic amber weevils.
Locality. Baltic amber.
Curculioninae incertae sedis
Protoceletes hirtus Nazarenko et Perkovsky, 2016
Nazarenko, Perkovsky [68]: 992, 994–995, Figure 1 therein, plate 7, Figure 1 therein.
Locality. Rovno amber.
Remarks. Holotype is kept in SIZK.
Entiminae Schoenherr, 1823
Key to tribes of Entiminae in Eocene amber
1. Mandibles without scar of deciduous process..................................................................................2
—Mandibles with scar of deciduous process........................................................................................3
2. Mandibles without scales. Rostrum usually long.............................................................. Hyperini
—Mandibles covered with scales. Rostrum short..................................................................[Sitonini]
3. Pronotum with postocular lobes........................................................................................................ 4
—Pronotum without postocular lobes...................................................................................................5
4. Mandibles with three long setae, bare. Humeri rounded. Maxillae covered laterally by prementum.......................................................................................................................Trachyphloeini, part
—Mandibles with more than three long setae, partially covered with scales. Humeri more or less convex. Maxillae exposed...................................................................................................Eudiagogini
5. Rostrum longer, distinctly longer than wide............................................................ Tropiphorini
—Rostrum short, at most slightly longer than wide...........................................................................6
6. Claws free.............................................................................................................................................7
—Claws connate at base...........................................................................................................................8
7. Apex of rostrum with carina forming posterior edge of large epistoma.................Anypotactini
—Posterior border of epistoma indistinct without carina.................................................Naupactini
8. Antennal scrobes dorsally in first half and laterally in second half of rostrum............................................................................................................................ Trachyphloeini, part.
—Antennal scrobes laterally................................................................................................ Polydrusini
Hyperini Marseul, 1863
Key to subtribes of Hyperini in Baltic amber
1. Mesepimeron widely trapezoidal. Metepisternum wide, expanded upward and downward. Prementum large................................................................................................................................ Cepurina
—Mesepimeron narrowly triangular. Metepisternum narrow, usually extended upward. Prementum small............................................................................................................................ [Hyperina]
Cepurina Capiomont, 1867
Genus Palaeophelypera Legalov, 2013
Type species: Palaeophelypera kuscheli Legalov, 2013
Palaeophelypera kuscheli Legalov, 2013 (Figure 10e,f)
Legalov [41]: 75–76, Figures 27–30 therein.
Locality. Baltic amber.
Remarks. This species is known only by the holotype (ISEA).
[Hyperina Marseul, 1863]
Remarks. The genus similar to Hypera Germar, 1817 was recorded from Baltic amber [61].
[Sitonini Gistel, 1856]
Tropiphorini Marseul, 1863
Key to genera of Tropiphorini in Baltic amber
1. Eyes strongly displaced downward. Elytra about 2.3 times as long as pronotum.....................................................................................................................................Scuccinalophus
—Eyes simple. Elytra 4.0 times as long as pronotum.........................................................Limalophus
Genus Limalophus Scudder, 1893
Type species: Limalophus compositus Scudder, 1893
Remarks. It is the first Old-World species of the genus Limalophus in Baltic amber.
urn:lsid:zoobank.org:act:9D578D48-EAA9-4375-BABB-84F4B2DEFDE2
Description. Size. Body length (without rostrum) 8.1 mm. rostrum length 1.6 mm. Body black, covered with quite narrow appressed scales. Head. Rostrum quite long, 1.3 times as long as pronotum, 4.1 times as long as wide in middle, weakly curved, densely punctate, with two longitudinal carinae. Scrobes distinct, lateral, slightly visible dorsally. Forehead quite wide, convex. Eyes small, convex. Vertex weakly flattened, punctate. Mandible massive, with scar. Antennae geniculate, inserted at apical third of rostrum laterally, with sparse semierect setae. Scape 6.4 times as long as wide, not reaching eye. Antennomeres: 2–8—conical; 2—1.4 times as long as wide, 0.3 times as long as and 1.3 times as wide as scape; 3—1.3 times as long as wide, 0.6 times as long as and 0.7 times as narrow as antennomere 2; 4—1.1 times as long as wide, equal in length and 1.2 times as wide as antennomere 3; 5—0.9 times as long as wide, 0.8 times as long as and equal to width to antennomere 4; 6—equal in length and width, 1.1 times as long as and equal in width to antennomere 5; 7—0.9 times as long as wide, equal in length and 1.1 times as wide as antennomere 6; 8—0.7 times as long as wide, subequal in length and 1.3 times as wide as antennomere 7. Club compact, 1.8 times as long as wide, 0.5 times as long as funicle, weakly acuminate. Pronotum bell-shaped, convex, densely punctate. Mesonotum. Scutellum triangular, small. Elytra elongate and distinctly convex, 4.0 times as long as pronotum. Humeri distinct. Striae regular and distinct. Interstriae convex, wide, 6.0–7.0 times as wide as width of striae, with dense appressed narrow scales and rarely decumbent scales. Thorax. Prosternum densely punctate, without postocular lobes. Precoxal part of prosternum 0.4 times as long as procoxal length. Procoxal cavities round, contiguous. Metaventrite long, 1.5 times as long as metacoxal length. Abdomen weakly convex. Ventrites oriented in one plane. Ventrite 1, 1.2 times as long as metacoxa. Ventrite 2, 0.8 times as long as ventrite 1. Ventrite 3, 0.7 times as long as ventrite 2. Ventrite 4, 0.8 times as long as ventrite 3. Ventrite 5, 1.6 times as long as ventrite 4. Legs elongate. Femora weakly clavate, without teeth. Profemora length/width ratio 3.0. Mesofemora length/width ratio 3.9. Metafemora length/width ratio 3.1. Tibiae almost straight, weakly flattened, without mucro and uncus. Metatibia length/width ratio 4.1. Tarsi quite long, with pulvilli on underside. Tarsomeres: 1 and 2—conical; 3—bilobed; 5—elongate. Claws large, free, without teeth.
Material examined. Holotype (ISEA), no. BA2017/6.
Comparison. The new species differs from L. contractus Scudder, 1893 from Green River in the longer and narrower rostrum, and larger body sizes.
Etymology. The epithet of this new species is dedicated to George Poinar, Jr. (Corvallis) who contributed to the studies of the amber ecosystems.
Locality. Baltic amber.
Remarks. The new species belongs to Entiminae based on the tibiae lacking an uncus and the massive mandible with a scar. The quite long rostrum, free tarsal claws and lateral antennal scrobes indicate that the new genus belongs to Tropiphorini. Simple eyes and quite short pronotum make it possible to place the new species in the genus Limalophus.
Genus Scuccinalophus Legalov, 2016
Type species: Scuccinalophus attenboroughi Legalov, 2016
Scuccinalophus attenboroughi Legalov, 2016 (Figure 11a)
Legalov [44]: 64–66, Figure 6 therein.
Locality. Baltic amber.
Remarks. This species is known only by the holotype (ISEA).
Eudiagogini LeConte, 1874
Genus Tolstonosik Legalov, Kirejtshuk et Nel, 2019
Type species: Tolstonosik oisensis Legalov, Kirejtshuk et Nel, 2019
Tolstonosik oisensis Legalov, Kirejtshuk et Nel, 2019
Legalov et al. [20]: 81–82, Figure 3c,f therein, plate 20, Figures 5 and 6 therein.
Locality. Oise amber.
Remarks. This species is known only by the holotype (MNHN).
Anypotactini Champion, 1903
Genus Paonaupactus Voss, 1953
Type species: Paonaupactus sitonitoides Voss, 1953
= Pareustolus Voss, 1953
Type species: Polydrosus scheelei Voss, 1953
= Protonaupactus Zherikhin, 1971, syn. nov.
Type species: Protonaupactus microphthalmus Zherikhin, 1971
= Sucinophyllobius Wanat et Borowiec, 1986
Type species: Sucinophyllobius viridis Wanat et Borowiec, 1986
Remarks. Small differences in the shape of the antennal club of species give not enough reasons to separate Protonaupactus into a separate genus.
Figure 11.
Habitus of Entiminae from Baltic and Rovno ambers. (a) Scuccinalophus attenboroughi, holotype, ISEA, no. BA2015/1, body, laterally. (b) Polydrosus scheelei, holotype, GPIH, no. 198, body, laterally. (c) Paonaupactus sitonitoides, holotype, GPIH, no. 197, body, ventrally. (d) Pyllobius cephalotes, holotype, ZMUC, no. 962, body, laterally. (e) Otiorhynchus pellucidipes, holotype, ZMUC, no. 961, body, laterally. (f) Paonaupactus sitonitoides, holotype, GPIH, no. 197, dorso-laterally; (g) P. viridis, holotype, EIW, no. 5637, body, laterally. (h,j) Arostropsis gusakovi sp. nov., holotype, CVGM, no. 032C2014: body, laterally (h); body, latero-ventrally (j). (i) Paonaupactus katyae, holotype, MAIG, no. 5981, body, laterally. Scale bars: 1.0 mm. See Section 2 for names of collections.
Figure 11.
Habitus of Entiminae from Baltic and Rovno ambers. (a) Scuccinalophus attenboroughi, holotype, ISEA, no. BA2015/1, body, laterally. (b) Polydrosus scheelei, holotype, GPIH, no. 198, body, laterally. (c) Paonaupactus sitonitoides, holotype, GPIH, no. 197, body, ventrally. (d) Pyllobius cephalotes, holotype, ZMUC, no. 962, body, laterally. (e) Otiorhynchus pellucidipes, holotype, ZMUC, no. 961, body, laterally. (f) Paonaupactus sitonitoides, holotype, GPIH, no. 197, dorso-laterally; (g) P. viridis, holotype, EIW, no. 5637, body, laterally. (h,j) Arostropsis gusakovi sp. nov., holotype, CVGM, no. 032C2014: body, laterally (h); body, latero-ventrally (j). (i) Paonaupactus katyae, holotype, MAIG, no. 5981, body, laterally. Scale bars: 1.0 mm. See Section 2 for names of collections.

Key species of genus Paonaupactus in Eocene amber
1. Scape barely extends beyond posterior margin of eye........................................................P. katyae
—Scape extends beyond front edge of eye and usually extends beyond front edge of pronotum............................................................................................................................................................2
2. Club oval. Elytral interstriae with 5–6 oval scales transverse to wide axis and sparse narrower scales...............................................................................................................................................P. sitonitoides
—Club fusiform.........................................................................................................................................3
3. Body covered with appressed hairs without scales.......................................... P. microphthalmus
—Body covered with scales..................................................................................................................... 4
4. Pronotum covered with wide scales...................................................................................P. gracilis
—Pronotum covered with narrow scales..............................................................................................5
5. Body (without rostrum) larger (5.2–6.2 mm). Apex of metatibiae simple in male........................................................................................................................................................ P. viridis
—Body (without rostrum) smaller (4.2–4.5 mm). Metatibiae strongly spatulate apically in male.................................................................................................................................................... P. sobrinus
Paonaupactus gracilis Legalov, Nazarenko et Perkovsky, 2019
Legalov et al. [72]: 73, 75, Figures 1f and 2b therein, plate 10.
Locality. Rovno amber.
Remarks. This species is known only by the holotype (SIZK). The length of its body (without rostrum) is 4.2 mm.
Paonaupactus katyae Legalov, Nazarenko and Perkovsky, 2019 (Figure 11i)
Legalov et al. [72]: 70–71, 73, Figures 1e and 2a therein, plate 9.
Locality. Rovno amber.
Remarks. This species is known only by the holotype (MAIG). The length of its body (without rostrum) is 3.6 mm.
Paonaupactus microphthalmus (Zherikhin, 1971)
Zherikhin [26]: 199, 205–206, Figure 3 therein, pl. X, Figure 2 therein (Protonaupactus).
Remarks. The holotype should be deposited in PIN; however, it was not found there. The length of its body (without rostrum) is 4.0 mm.
Paonaupactus sitonitoides Voss, 1953 (Figure 11b–f)
Voss [24]: 128, Figure 6 therein.
=Polydrosus scheelei Voss [24]: 129–130, Figure 8 therein.
=Pyllobius cephalotes Voss [25]: 175–176, Figure 6 therein.
=Otiorhynchus pellucidipes Voss [25]: 173, Figure 4 therein.
Locality. Baltic amber.
Remarks. Holotypes of P. sitonitoides (Figure 11c,f) and Polydrosus scheelei (Figure 11b) are kept in the GPIH, and Pyllobius cephalotes (Figure 11d) and Otiorhynchus pellucidipes (Figure 11e) are kept in the SZUC. The length of their body (without rostrum) varies within 3.7–4.8 mm.
Paonaupactus sobrinus (Voss, 1972)
Voss [25]: 174–175, Figures 5 and 12 therein (Phyllobius).
Locality. Baltic amber.
Remarks. Holotype is kept in the SZUC. The length of body (without rostrum) is 4.5 mm.
Paonaupactus viridis (Wanat et Borowiec, 1986) (Figure 11g,f)
Wanat, Borowiec [30]: 244–246, Figures 1–3 therein (Sucinophyllobius)
Locality. Baltic amber.
Remarks. Length of body (without rostrum) 5.2–6.2 mm. Holotype is kept in the EIW and one specimen (CVGM), no. 031C2000 was studied. The length of the body of the mentioned specimens (without rostrum) varies from 5.2 to 6.2 mm.
Naupactini Gistel, 1856
Genus Arostropsis Yunakov et Kirejtshuk, 2011
Type species: Arostropsis groehni Yunakov et Kirejtshuk, 2011
Key to species of genus Arostropsis in Eocene amber
1. Antennomere 2 subequal to antennomere 3. Elytra wider with weaker convex humeral calli. Pronotum wider than long........................................................................................................... A. perkovskyi
—Antennomere 2 distinctly longer than antennomere 3. Elytra narrower with distinctly convex humeral calli......................................................................................................................................................2
2. Eyes oval. Pronotum distinctly narrower than long in middle.......................................A. groehni
—Eyes rounded. Pronotum subequal in length and width in middle...............A. gusakovi sp. nov.
Arostropsis groehni Yunakov et Kirejtshuk, 2011
Yunakov, Kirejtshuk [37]: 6–10, Figures 1–16 therein.
Locality. Baltic amber.
Remarks. This species is known only by the holotype (GPIHG).
Arostropsis gusakovi sp. nov. (Figure 11h,j)
urn:lsid:zoobank.org:act:E9215A77-7228-4D07-95BE-CE26E465F9D6
Description. Female. Size. Body length (without rostrum) 11.5 mm. rostrum length 1.5 mm. Body black–brown, covered with dense rounded green appressed scales. Head. Rostrum quite short, 0.7 times as long as pronotum, 1.5 times as long as wide in middle, punctate, with longitudinal line not reaching vertex. Posterior border of epistoma indistinct, without carina. Scrobes laterally. Forehead wide, flattened. Eyes small, rounded, convex. Vertex weakly flattened, densely punctate. Temples longer than eyes. Antennae geniculate, inserted behind middle of rostrum. Scape 6.2 times as long as wide, extends beyond front edge of eye. Antennomeres: 2–8—long, conical; 2—3.0 times as long as wide, 0.4 times as long as and 0.9 times as narrow as scape; 3—2.2 times as long as wide, 0.7 times as long as and 0.9 times as narrow as antennomere 2; 5—1.7 times as long as wide; 5–7—equal in width; 6—2.0 times as long as wide, 1.2 times as long antennomere 5; 7—equal to sixth; 8—1.4 times as long as wide, 0.8 times as long as and 1.2 times as wide as antennomere 7. Club compact, 2.5 times as long as wide, weakly acuminate. Pronotum bell-shaped, 1.1 times as long as wide apically, subequal in length and width medially, 0.9 times as long as wide basally, weakly flattened, finely and densely punctate. Mesonotum. Scutellum triangular, small. Elytra elongate and distinctly convex, 3.1 times as long as pronotum, 2.3 times as long as wide at base, 1.8 times as long as wide in middle, 2.5 times as long as wide in apical quarter. Humeri weakly smoothed. Striae regular and distinct. Interstriae flat, wide, 4.3–5.7 times as wide as width of striae. Thorax. Prosternum densely punctate, without postocular lobes. Precoxal part of prosternum 0.4 times as long as procoxal length. Procoxal cavities round, contiguous. Postcoxal part of prosternum 0.3 times as long as procoxal length. Metaventrite long, 3.0 times as long as metacoxal length. Metepisternum 6.6 times as long as wide in middle. Abdomen weakly convex. Ventrites oriented in one plane. Ventrite 1, 1.4 times as long as metacoxa. Ventrite 2, 1.1 times as long as ventrite 1. Ventrite 3, 0.6 times as long as ventrite 2. Ventrite 4, 0.7 times as long as ventrite 3. Ventrite 5, 2.2 times as long as ventrite 4. Legs elongate. Femora weakly clavate, without teeth. Profemora length/width ratio 3.6. Mesofemora length/width ratio 4.1. Metafemora length/width ratio 3.3. Tibiae almost straight, weakly flattened, without mucro and uncus. Metatibial corbels open. Protibia length/width ratio 7.6. Mesotibia length/width ratio 5.6. Metatibia length/width ratio 7.1. Tarsi quite long, with pulvilli on underside. Tarsomeres: 1 and 2—conical; 3—bilobed; 5—elongate. Claws large, free, without teeth. Protarsomeres: 1—1.7 times as long as wide; 2—equal in length and width, 0.7 times as long as and 1.1 times as wide as first; 3—0.6 times as long as wide, equal in length and 1.8 times as wide as second; 5—3.3 times as long as wide, 1.3 times as long as and 0.2 times as narrow as third; mesotarsomeres: 1—1.7 times as long as wide; 2—equal in length and width, 0.6 times as long as and 1.1 times as wide as tarsomere 1; 3—0.6 times as long as wide, equal in length and 1.6 times as wide as tarsomere 2; 5—5.0 times as long as wide, 1.9 times as long as and 0.2 times as narrow as tarsomere 3; metatarsomeres: 1—1.5 times as long as wide; 2—1.3 times as long as wide, 0.8 times as long as and equal in width to tarsomere 1; 3—0.6 times as long as wide, 0.8 times as long as and 1.6 times as wide as tarsomere 2; 5—5.0 times as long as wide, 1.9 times as long as and 0.2 times as narrow as tarsomere 3.
Material examined. Holotype (CVGM), no. 032C2014.
Comparison. The new species differs from A. groehni in the rounded eyes, pronotum with subequal length and width in middle and larger body sizes.
Etymology. The epithet of this new species is dedicated to Viktor A. Gusakov (Moscow).
Locality. Baltic amber.
Arostropsis perkovskyi Bukejs et Legalov, 2019
Bukejs, Legalov [71]: 26, Figures 1 and 2 therein.
Locality. Rovno amber.
Remarks. This species is known only by the holotype (GPIHG).
Polydrusini Schoenherr, 1823
Genus Polydrusus Germar, 1817
Type species: Curculio undatus Fabricius, 1781 (= Curculio tereticollis DeGeer, 1775)
Subgenus Palaeodrosus Zherikhin, 1971
Type species: Polydrusus archetypus Zherikhin, 1971
Key to species of subgenus Palaeodrosus in Baltic amber
1. Elytra narrower. Pronotum and elytra covered with narrow scales. Eyes slightly convex...................................................................................................................................... P. (P.) archetypus
—Elytra wider. Pronotum covered with wide scales. Elytra covered with two types of scales: narrow erect and wide semierect. Eyes more convex.......................................... P. (P.) zherikhini sp. nov.
Polydrusus (Palaeodrosus) archetypus Zherikhin, 1971
Zherikhin [26]: 206–207, Figure 4 therein.
Locality. Baltic amber.
Remarks. The holotype of this species should be deposited in PIN; however, at the moment, it is missing there.
urn:lsid:zoobank.org:act:FB4E96D8-3069-4D91-86E1-C8950252441D
Description. Female. Size. Body length (without rostrum) 2.9 mm. rostrum length 0.4 mm. Body black, covered with sparse narrow and wide scales. Head. Rostrum quite short, 0.7 times as long as pronotum, 0.9 times as long as wide in middle, 0.8 times as long at base, punctate. Scrobes lateral. Forehead wide, flattened, punctate. Eyes large, rounded, convex. Vertex weakly flattened. Temples 0.5 times as long as eyes. Antennae geniculate, inserted behind middle of rostrum. Scape 6.7 times as long as wide, not reaching pronotum. Antennomeres: 2–8—conical; 2—1.7 times as long as wide, 0.3 times as long as and subequal in width to scape; 3—2.0 times as long as wide, 0.6 times as long as and 0.5 times as narrow as antennomere 2; 4—1.4 times as long as wide, 0.8 times as long as and 1.2 times as wide as antennomere 3; 5—equal to antennomere 4. Club compact, quite long, weakly acuminate. Pronotum bell-shaped, 0.9 times as long as wide apically, 0.6 times as long as wide medially, 0.7 times as long as wide basally, weakly flattened, densely punctate, covered with wide scales. Sides weakly rounded. Mesonotum. Scutellum suboval, small. Elytra convex, quite wide, 3.7 times as long as pronotum, 1.7 times as long as wide at base, 1.5 times as long as wide in middle, 2.6 times as long as wide in apical quarter. Humeri weakly smoothed. Striae regular and distinct. Interstriae flat, wide, 1.5–2.0 times as wide as width of striae, with narrow erect, and semierect wide scales. Thorax. Prosternum punctate, without postocular lobes. Pre- and postcoxal parts of prosternum short. Procoxal cavities round, contiguous. Metaventrite equal in length to metacoxal length. Metepisternum 5.1 times as long as wide in middle. Abdomen convex. Ventrite 1, 1.4 times as long as metacoxa. Ventrite 2, 1.1 times as long as ventrite 1. Ventrite 3, 0.4 times as long as ventrite 2. Ventrite 4, 0.8 times as long as ventrite 3. Ventrite 5, 2.8 times as long as ventrite 4. Legs elongate. Femora weakly clavate, without teeth. Tibiae almost straight, weakly flattened, without mucro and uncus. Metatibial corbels open. Tarsi quite long, with pulvilli on underside. Tarsomeres: 1 and 2—conical; 3—bilobed; 5—elongate. Claws large, fused at base, without teeth.
Material examined. Holotype (ISEA), BA2015/14.
Comparison. The new species differs from P. (P.) archetypus Zherikhin, 1971 in the wider elytra, pronotum covered with wide scales, elytra with narrow erect, and semierect wide scales and also in the more convex eyes.
Etymology. The epithet of this new species is dedicated to the paleoentomologist Vladimir V. Zherikhin.
Locality. Baltic amber.
Figure 12.
Habitus of Entiminae and Platypodidae from Baltic amber. (a) Polydrusus zherikhini sp. nov., holotype, ISEA, BA2015/14, body, laterally. (b,e) Archaeocallirhopalus larssoni, holotype, ISEA, no. BA2012/1: body, dorsally (b); body, laterally (e). (c,f) Archaeosciaphilus marshalli, holotype, EIW, no. 7975: body, laterally, on the left (c); body, laterally, on the right (f). (d) Polydrusus zherikhini sp. nov., holotype, ISEA, BA2015/14, body, laterally. (g) Archaeocallirhopalus alekseevi, paratype, ISEA, no. BA2012/13, body, laterally (g). (h,i) Cenocephalus aniskini sp. nov., holotype, ISEA, no. BA2018/3: body, dorsally (h); body, ventrally (i). Scale bars: 1.0 mm. See Section 2 for names of collections.
Figure 12.
Habitus of Entiminae and Platypodidae from Baltic amber. (a) Polydrusus zherikhini sp. nov., holotype, ISEA, BA2015/14, body, laterally. (b,e) Archaeocallirhopalus larssoni, holotype, ISEA, no. BA2012/1: body, dorsally (b); body, laterally (e). (c,f) Archaeosciaphilus marshalli, holotype, EIW, no. 7975: body, laterally, on the left (c); body, laterally, on the right (f). (d) Polydrusus zherikhini sp. nov., holotype, ISEA, BA2015/14, body, laterally. (g) Archaeocallirhopalus alekseevi, paratype, ISEA, no. BA2012/13, body, laterally (g). (h,i) Cenocephalus aniskini sp. nov., holotype, ISEA, no. BA2018/3: body, dorsally (h); body, ventrally (i). Scale bars: 1.0 mm. See Section 2 for names of collections.

Genus Archaeosciaphilus Legalov, 2012
Type species: Archaeosciaphilus marshalli Legalov, 2012
Remarks. This genus was placed in Sciaphilini Sharp, 1891. The study of an undescribed species of the genus with clearly visible characters showed that it is characterized by mandibles with five long setae and maxillae exposed at the sides of the prementum, making it possible to transfer it to Polydrusini [Legalov, Poinar, in preparation].
Archaeosciaphilus marshalli Legalov, 2012 (Figure 12c,f)
Legalov [39]: 271, Figure 2d therein, pl. 9, Figure 2 therein.
Locality. Baltic amber.
Remarks. This species is known by the holotype (EIW).
Trachyphloeini Lacordaire, 1863
[Trachyphloeina Lacordaire, 1863]
Remarks. The genus Trachyphloeus Germar, 1817 was recorded from Baltic amber [61].
Pseudocneorrhinina Kono, 1930
Genus Archaeocallirhopalus Legalov, 2013
Type species: Archaeocallirhopalus larssoni Legalov, 2013
Key species of genus Archaeocallirhopalus in Baltic amber
1. Body (without rostrum) larger (5.1 mm). Forehead with strong tubercles over eyes. Scape 4.1 times as long as wide......................................................................................................................... A. larssoni
—Body (without rostrum) smaller (3.3 mm). Forehead without strong tubercles over eyes. Scape 3.7 times as long as wide...................................................................................................... A. alekseevi
Archaeocallirhopalus alekseevi Legalov et Bukejs, 2015 (Figure 12g)
Legalov, Bukejs [43]: 25–28, Figures 1–5 therein.
Locality. Baltic amber.
Remarks. This species is known by the holotype (ABCD) and the paratype (ISEA).
Archaeocallirhopalus larssoni Legalov, 2013 (Figure 12b,e)
Legalov [41]: 77–78, Figures 31–34 therein.
Locality. Baltic amber.
Remarks. The holotype is kept in ISEA.
Curculionidae incertae sedis
Genus Thryogenosoma Voss, 1953
=Erirhinoides Motschulsky, 1857 non Blanchard, 1849
Type species: Erirhinoides cariniger Motschulsky, 1857
Thryogenosoma cariniger (Motschulsky, 1857)
Motschulsky [22]: 27 (Erirhinoides).
Locality. Baltic amber.
Remarks. The beetle was deposited in the Danziger Naturkundemuseum Menge collection which was lost during the Second World War.
Platypodidae Shuckard, 1839
Tesserocerinae Strohmeyer, 1914
Tesserocerini Strohmeyer, 1914
Key to genera of Tesserocerini in Baltic amber
1. Mesonotum without longitudinal carina. Antennal club wider than long................. Eoplatypus
—Mesonotum with longitudinal carina. Antennal club longer than wide.................Cenocephalus
Genus Eoplatypus Cognato et Smith, 2019
Type species: Eoplatypus jordali Peris, Solórzano Kraemer et Cognato, 2017
Eoplatypus jordali Peris, Solórzano Kraemer et Cognato, 2017
Peris et al. [93]: 191, Figure 4 therein.
Locality. Baltic amber.
Remarks. The species is known by the holotype and paratype from SFNF [93].
Genus Cenocephalus Chapuis, 1865
Type species: Cenocephalus thoracicus Chapuis, 1865
Remarks. This find is the first record of the genus in Baltic amber.
Cenocephalus aniskini sp. nov. (Figure 12h,i)
urn:lsid:zoobank.org:act:4B9EDF95-BCCD-4F3A-BF1E-DA4E4550A9F9
Description. Description. Female. Size. Body length (without rostrum) 3.2 mm. Body yellow–brown, naked. Head. Head subequal in width to pronotum, ventrally with pregular sutures. Forehead wide, weakly concave, with abundant, long, erect setae. Eyes large, weakly convex, rounded, coarsely faceted. Temples much shorter than eye. Rostral pleurostomal sinus shallow. Maxilla with mesal clement clearly divided into separate lacinia and galea. Scape elongate, reaching second third of eye, 3.8 times as long as wide. Antennomeres: 2–5—wide, conical; 2—0.7 times as long as wide, 0.01 times as long as and 0.5 times as narrow as scape; 3—0.6 times as long as wide, 1.5 times as long as and 1.7 times as wide as antennomere 2; 4—0.3 times as long as wide, 0.7 times as long as and 1.1 times as wide as antennomere 3; 5—about 0.3 times as long as wide, equal in length and 1.1 times as wide as fourth. Club compact, flat, 1.5 times as long as wide, 2.5 times as long as funicle, without sutures between segments of club. Pronotum subcylindrical, 1.4 times as long as wide apically, 1.3 times as long as wide medially and 1.4 times as long as wide basally, finely punctate, weakly convex. Sides straight, parallel. Disc with minute punctures. Posterior one-third of pronotum with transverse band of numerous, small, closely placed mycetangia pores. Mesonotum with longitudinal carina. Scutellum almost triangular, small, located below surface of elytra. Elytra elongate, 1.8 times as long as pronotum, 2.6 times as long as wide basally, 2.4 times as long as wide in middle, 2.5 times as long as wide in apical quarter. Elytral base carinate. Humeri smoothed. Striae weak. Interstriae flat, wide, 4.0–5.0 times as wide as striae. Elytral declivity commencing in apical fourth, slightly concave, with erect setae, without tubercles. Border of declivity armed with two distinct long spines on interstria 3. Thorax. Prothorax finely punctate. Posterior margin of prothorax strongly procurved in pleural area. Femoral grooves distinct. Precoxal part of prosternum 0.7 times as long as length of procoxa. Procoxal cavities contiguous. Metaventrite flat, finely punctate, 3.9 times as long as metacoxal length. Metepisternum quite wide, 3.7 times as long as wide in middle. Abdomen weakly convex. Ventrite 1, 0.4 times as long as metacoxal length. Ventrite 2, equal in length to ventrite 1. Ventrite 3, 2.2 times as long as ventrite 2. Ventrite 4, 1.2 times as long as ventrite 3. Ventrite 5, equal in length to ventrite 4. Legs elongate. Procoxae enlarged. Mesocoxae rounded, widely separated. Femora laterally compressed, wide. Metafemora 2.2 times as long as wide in middle. Tibiae curved, with three lateral ridges or rugae on outer surfaces, with mucro. Metatibia 2.7 times as long as wide in middle. Tarsi elongate. Tarsomeres: 1—long, longer than tarsomeres 2–5 combined; 2 and 3—long, conical; 4—slightly longer than wide; 5—elongate. Claws free and long.
Material examined. Holotype (ISEA), no. BA2018/3.
Comparison. The new species differs from other species of the genus in the border of the elytral declivity armed with two, distinct, long spines on the interstria 3 and slightly concave unarmed elytral declivity.
Etymology. The epithet of this new species is dedicated to Dr. V. M. Aniskin (Khristianovich Institute of Theoretical and Applied Mechanics, SB RAS, Novosibirsk) who helped the author with the study.
Locality. Baltic amber.
Remarks. The new species belongs to Tesserocerinae because the maxilla with mesal element is clearly divided into separate lacinia and galea. Contiguous procoxal cavities give a reason to put this new species in tribe Tesserocerini. Mesonotum with longitudinal carina, antennal club longer than wide and rounded eyes make it possible to place it in the genus Cenocephalus.
Spectra of laser-induced fluorescence (LIF) were investigated to confirm the belonging of this amber sample.
Scolytidae Latreille, 1804
Key subfamilies of Scolytidae in Eocene amber
1. Head visible dorsally. Anterior margins of elytra procurved and with rows of crenulations....................................................................................................................................Hylesininae
—Head concealed dorsally by pronotum. Anterior margins of elytra forming straight line, unarmed............................................................................................................................................. Scolytinae
Hylesininae Erichson, 1836
Key to tribes of Hylesininae in Eocene amber
1. Precoxal portion of prosternum strongly elevated.............................................................Hylastini
—Precoxal portion of prosternum not elevated................................................................................... 2
2. Eyes simple, not emarginate............................................................................................... Hylurgini
—Eyes distinctly emarginate...................................................................................................................3
3. Antennal club symmetrical, sutures transverse......................................................... Polygraphini
—Antennal club with three oblique sutures..................................................................... Phloeosinini
Hylastini LeConte, 1876
Genus Hylastes Erichson, 1836
Type species: Bostrichus ater Paykull, 1800
Hylastes aterites Schedl, 1947
Schedl [23]: 21–22.
Locality. Baltic amber.
Remarks. This species is known by two syntypes [23], which should be deposited in the collection of the Albertus-Universität Königsberg and which was lost during the Second World War.
Genus Hylurgops LeConte, 1876
Type species: Hylastes pinifex Fitch, 1858 (= Hylastes rugipennis Mannerheim, 1843)
= Hylesinites Germar, 1813
Type species: Hylesinites electrinus Germar, 1813
= Hylastites Hagedorn, 1906
Type species: Hylastites schellwieni Hagedorn, 1906
= Myelophilites Hagedorn, 1907
Type species: Myelophilites dubius Hagedorn, 1906
= Hylescierites Schedl, 1947
Type species: Hylescierites granulatus Schedl, 1947
Remarks. The key to species is based on the key from Schedl [23]. H. electrinus and H. schellwieni are not included in the key because it requires a study of the types.
Key species of genus Hylurgops in Baltic amber
1. Procoxal cavities distinctly separated. Body smaller (2.4 mm)................................. H. granulatus
—Procoxal cavities almost contiguous. Body larger (2.9–3.5 mm).....................................................2
2. Body smaller (2.9 mm)........................................................................................................... H. dubius
—Body larger (3.2–3.5 mm)......................................................................................................................3
3. Body very slender. Pronotum distinctly narrowed to apex........................................ H. pilosellus
—Body stumpy, more cylindrical. Pronotum weakly narrowed to apex..........................................4
4. Elytral declivity punctate.............................................................................................. H. corpulentus
—Elytral declivity with granules. Each granule with long setae................................ H. tuberculatus
Hylurgops corpulentus Schedl, 1947
Schedl [23]: 23, 27–28, Figures 10 and 11 therein.
Locality. Baltic amber.
Remarks. This species is known by a series of syntypes [23], which should be deposited in the collection of the Albertus-Universität Königsberg and which was lost during the Second World War.
Hylurgops dubius (Hagedorn, 1906)
Hagedorn [84]: 118 (Myelophilites).
Locality. Baltic amber.
Remarks. This species is known by the holotype and a specimen [23], which should be deposited in the collection of the Albertus-Universität Königsberg and which was lost during the Second World War.
Hylurgops electrinus (Germar, 1813)
Germar [21]: 15 (Hylesinites).
Locality. Baltic amber.
Remarks. The holotype is kept in the MLUH. Description without any information about reposition of the specimen examined.
Hylurgops granulatus (Schedl, 1947)
Schedl [23]: 30, Figure 12 therein (Hylescierites).
Locality. Baltic amber.
Remarks. This species is known by the holotype [23], which should be deposited in the collection of the Albertus-Universität Königsberg and which was lost during the Second World War.
Hylurgops pilosellus Schedl, 1947
Schedl [23]: 23, 26–27, Figure 9 therein.
Locality. Baltic amber.
Remarks. This species is known by a series of syntypes [23], which should be deposited in the collection of the Albertus-Universität Königsberg and which was lost during the Second World War.
Hylurgops schellwieni (Hagedorn, 1906)
Hagedorn [94]: 117 (Hylastites).
Locality. Baltic amber.
Remarks. The holotype was lost [23].
Hylurgops tuberculatus Schedl, 1947
Schedl [23]: 23, 28–29.
Locality. Baltic amber.
Remarks. This species is known by a series of syntypes [23], which should be deposited in the collection of the Albertus-Universität Königsberg and which was lost during the Second World War.
Hylurgini Gistel, 1848
Key to genera of Hylurgini in Eocene amber
1. Procoxae very narrowly separated.................................................................................. Xylechinites
—Procoxae widely separated.................................................................................................................. 2
2. Club conical, with two transverse sutures. Its apical segment pointed........................ Xylechinus
—Club flattened, wide, with three distinctly transverse sutures. Its apical segment rounded.................................................................................................................................................. Klesovia
Genus Xylechinus Chapuis, 1869
Type species: Hylesinus pilosus Ratzeburg, 1837
Xylechinus mozolevskae Petrov et Perkovsky, 2008
Petrov, Perkovsky [65]: 407, Figure 1 therein.
Locality. Rovno amber.
Remarks. This species is known by the holotype in SIZK [65].
Genus Klesovia Petrov et Perkovsky, 2018
Type species: Klesovia pubescens Petrov et Perkovsky, 2018
Klesovia pubescens Petrov et Perkovsky, 2018
Petrov, Perkovsky [69]: 167, Figure 1 therein, plate 7, Figures 1 and 2 therein.
Locality. Rovno amber.
Remarks. This species is known by the holotype and several paratypes in SIZK [69].
Genus Xylechinites Hagedorn, 1906
Type species: Xylechinites anceps Hagedorn, 1906
Xylechinites anceps Hagedorn, 1906
Hagedorn [94]: 120, Figures 10–12 therein.
Locality. Baltic amber.
Remarks. This species is known by the holotype [23], which should be deposited in the collection of the Albertus-Universität Königsberg and which was lost during the Second World War.
Phloeosinini Nuesslin, 1912
Genus Phloeosinus Chapius, 1869
Type species: Hylesinus thujae Perris, 1855
= Phloeosinites Hagedorn, 1906
Type species: Phloeosinites rehi Hagedorn, 1906
Remarks. The key to species is based on the key from Schedl [23]. Ph. rehi is not included in the key, because its differences from other species need to be clarified.
Key species of genus Phloeosinus in Baltic amber
1. Elytral declivity without granules or teeth.......................................................................... Ph. wolffi
—Elytral declivity with granules or teeth..............................................................................................2
2. Elytral declivity with large teeth................................................................................. Ph. sexspinosus
—Elytral declivity with rows of granules on first and third interstriae.............................................3
3. Body slender, cylindrical.................................................................................................. Ph. robustus
—Body stumpy, oval.................................................................................................................................4
4. Elytral striae weakly visible. Elytral interstriae densely irregular punctate and with two weak, irregular rows of points.....................................................................................................................................5
—Elytral striae distinct. Elytral interstriae only with two weak, irregular rows of points.............6
5. Elytral declivity oblique and weakly convex, with very small granules............... Ph. tuberculifer
—Elytral declivity distinctly convex, with distinct rows of granules............................. Ph. assimilis
6. Interstriae of elytral declivity narrow, with sparse, quite large granules..................... Ph. brunni
—Interstriae of elytral declivity wide, with dense, small granules........................ Ph. regimontanus
Phloeosinus assimilis (Schedl, 1947)
Schedl [23]: 34, 37–38 (Phloeosinites).
Locality. Baltic amber.
Remarks. This species is known by the holotype [23], which should be deposited in the collection of the Albertus-Universität Königsberg and which was lost during the Second World War.
Phloeosinus brunni (Hagedorn, 1906)
Hagedorn [94]: 119, Figures 7 and 8 therein (Phloeosinites).
Locality. Baltic amber.
Remarks. This species is known by the holotype and a specimen [23], which should be deposited in the collection of the Albertus-Universität Königsberg and which was lost during the Second World War.
Phloeosinus regimontanus (Hagedorn, 1906)
Hagedorn [94]: 119–120, Figure 9 therein (Phloeosinites).
Locality. Baltic amber.
Remarks. This species is known by a series of syntypes [23], which should be deposited in the collection of the Albertus-Universität Königsberg and which was lost during the Second World War.
Phloeosinus rehi (Hagedorn, 1906)
Hagedorn [94]: 118 (Phloeosinites).
Locality. Baltic amber.
Remarks. This species is known by the holotype [94], which should be deposited in the collection of the Albertus-Universität Königsberg and which was lost during the Second World War.
Phloeosinus robustus (Schedl, 1947)
Schedl [23]: 34, 36–37 (Phloeosinites).
Locality. Baltic amber.
Remarks. This species is known by a series of syntypes [23], which should be deposited in the collection of the Albertus-Universität Königsberg and which was lost during the Second World War.
Phloeosinus sexspinosus (Schedl, 1947)
Schedl [23]: 34–36 (Phloeosinites).
Locality. Baltic amber.
Remarks. This species is known by the holotype [23], which should be deposited in the collection of the Albertus-Universität Königsberg and which was lost during the Second World War.
Phloeosinus tuberculifer (Schedl, 1947)
Schedl [23]: 34, 37 (Phloeosinites).
Locality. Baltic amber.
Remarks. This species is known by the holotype [23], which should be deposited in the collection of the Albertus-Universität Königsberg and which was lost during the Second World War.
Phloeosinus wolffi (Schedl, 1947)
Schedl [23]: 35, 40–41, Figure 16 therein (Phloeosinites).
Locality. Baltic amber.
Remarks. This species is known by the holotype [23], which should be deposited in the collection of the Albertus-Universität Königsberg and which was lost during the Second World War.
Polygraphini Chapuis, 1869
Genus Carphoborus Eichhoff, 1864
Type species: Hylesinus minimus Fabricius, 1801
=Carphoborites Carpenter, 1992
Type species: Charphoborites keilbachi Schedl, 1947
Key to species of genus Carphoborus in Baltic amber
1. Forehead convex. Body slender......................................................................................... C. keilbachi
—Forehead flat. Body stumpy................................................................................................ C. posticus
Carphoborus keilbachi (Schedl, 1947)
Schedl [23]: 32–33 (Carphoborites).
Locality. Baltic amber.
Remarks. This species is known by the holotype [23], which should be deposited in the collection of the Albertus-Universität Königsberg and which was lost during the Second World War.
Carphoborus posticus (Schedl, 1947)
Schedl [23]: 33 (Carphoborites).
Locality. Baltic amber.
Remarks. This species is known by the holotype [23], which should be deposited in the collection of the Albertus-Universität Königsberg and which was lost during the Second World War.
Scolytinae Latreille, 1804
Dryocoetini Lindemann, 1877
Key to genera of Dryocoetini in Eocene amber
1. Pronotum evenly convex................................................................................................... Taphramites
—Pronotum strongly convex in middle, flattened basally............................................. Taphrorychus
Genus Taphramites Schedl, 1947
Type species: Taphramites gnathotrichus Schedl, 1947
Key to species of genus Taphramites in Eocene amber
1. Elytral declivity with long, dense, thick setae......................................................... T. gnathotrichus
—Elytral declivity with short, slender setae...................................................................... T. rovnoensis
Taphramites gnathotrichus Schedl, 1947
Schedl [23]: 42.
Locality. Baltic amber.
Remarks. This species is known by the holotype [23], which should be deposited in the collection of the Albertus-Universität Königsberg and which was lost during the Second World War. This is a common species in Baltic amber [65].
Taphramites rovnoensis Petrov et Perkovsky, 2008
Petrov, Perkovsky [65]: 408, Figures 2–4 therein.
Locality. Rovno amber.
Remarks. This species is known by the holotype and paratypes in SIZK [65].
Genus Taphrorychus Eichhoff, 1878
Type species: Bostrichus bicolor Herbst, 1793
Taphrorychus immaturatus Schedl, 1947
Schedl [23]: 17, 43, 44, Figure 17 therein.
Locality. Baltic amber.
Remarks. This species is known by the holotype [23], which should be deposited in the collection of the Albertus-Universität Königsberg and which was lost during the Second World War.
4. Discussion
In total, 142 species of Curculionoidea [2,3,84,85,86,87,88] are recognized in this review, including nine species of Belidae, 10 species of Rhynchitidae, 13 species of Brentidae, 70 species of Curcuionidae, two species of Platypodidae, and 24 species of Scolytidae (Table 1; Figure 13). Oise amber has eight species, Baltic amber has 118 species, and Rovno amber has 16 species.
The Nemonychidae, a small family comprising the oldest weevils [12,84,95,96], is rare in amber [2,97,98,99,100]. Nemonychidae in Baltic amber is represented by one specialized species of a monotypic genus and tribe, which is characterized by more advanced features compared to modern representatives of Cimberidinae. Kuschelomacer kerneggeri is only present in Baltic amber and probably developed on Pinus like the recent species of this subfamily [96,101,102].
Figure 13.
Composition of the species in fauna of Eocene amber.

The Anthribidae is quite rare in the fossil record, although it is diverse in modern tropics and subtropics [103]. Representatives of all three modern subfamilies are found in Eocene amber. The Urodontinae was recorded only for Baltic amber [62], but the specimen requires re-study. The Choraginae is found in Rovno amber. The island tribe Valenfriesiini is mainly represented by a monotypic genus. There is no doubt that Choraginae will be found in Baltic amber also. The Anthribinae is found in all Eocene ambers. The monotypic tribe Oiserhinini is described from Oise amber. The genus Glaesotropis from Zygaenodini in Late Eocene amber is the richest in species (10). One species of this genus was found in Rovno amber. Nine species of three subgenera of this genus are described from Baltic amber. G. minor is one of the most common amber Curculionoidea. The Mecocerini, Allandrini, and possibly Tropiderini are noted only in Baltic amber.
The Belidae is a relict group [5,104]. The Belinae are not found in amber. It is possible that, in the Eocene, it was absent in Europe. The Oxycoryninae is represented by several species of the modern genus Oxycraspedus Kuschel, 1955 (tribe Oxycraspedini) and two genera of Metrioxenini. Species of the genus Oxycraspedus that develop on Araucaria [105] are not yet discovered in Baltic amber. Succinometrioxena poinari is one of the common Curculionoidea of Baltic amber. Probably, like modern Metrioxenini [105], fossil species developed on palm trees. The Belidae only appears in Baltic amber.
The Rhynchitidae is rare in the fossil record [2]. The Sayrevilleinae is represented by Sanyrevilleini with five species of the genus Baltocar. It is absent in French and Rovno ambers. Two tribes, Auletini and Rhynchitini, are found in Late Eocene amber. The modern genus Pseudomesauletes, which is known from the Oriental region, Africa, Madagascar, North and Central America, and the terminal Eocene of USA [106], is represented by one species in Rovno amber. Four species from four extinct genera are also described in Baltic amber. The genus Electrauletes is similar to the modern Mediterranean genus Auletes Schoenherr, 1826 and probably also developed on Cupressaceae [107]. The genus Eoropseudauletes belongs to Pseudauletina of Auletinini that is now widespread only in the Neotropics [106]. Representatives of the advanced tribes Deporaini and Byctiscini were not found in the fossil state.
The Brentidae is represented in the Eocene ambers by the subfamilies Apioninae and Nanophyinae. The Apioninae is found in Oise and Baltic ambers. The genus Palaeotanaos with one species from the monotypic tribe Palaeotanaini is described from Oise amber, where it is the most common of the Curculionoidea [20]. The Baltic amber Apioninae fauna is quite rich and represented by 11 species from eight genera of Rhadinocybini, Notapionini, Aspidapiini, Kalcapiini and Piezotrachelini. The genera Pseudaspidapion, Melanapion and Conapium are represented in the modern fauna. The Nanophyinae was found only in Baltic amber, where it is represented by one primitive genus. The remaining subfamilies of the brentid family are absent in amber, although Brentinae is recorded from the Middle Eocene deposits of Germany [107,108,109,110].
The Curculionidae is the largest family of weevils in the modern fauna and also the largest family found in amber. Eight subfamilies (Erirhininae, Molytinae, Lixinae, Dryophthorinae, Cossoninae, Conoderinae, Curculioninae, and Entiminae) are found in Eocene amber (Figure 14). The Cyclominae is extremely rare in the fossil record [2] and is not found in amber. Unfortunately, 13 tribes, from 40 tribes, noted in Baltic amber are known only from records [57,61] and their presence in amber requires confirmation. The Erirhininae is represented by Erirhinini, Dorytomini, and Bagoini. The most diverse species of the genus Dorytomus, which are associated with Salicaceae, are found in Late Eocene amber. The Molytinae has nine tribes. These are forms confined to wood or plant litter. In Rovno amber, only Cryptorhynchini is noted, which is also found in Oise and Baltic amber. The Acicnemidini, Pissodini, Magdalinini, Molytini, Plinthini, Mecysolobini, and Aedemonini are known only in Baltic amber. The genus Electrotribus, one of the most widespread genera in Baltic amber, is absent in other ambers. The Lixinae is known only by record of the genus Lixus in Baltic amber. The Dryophthorinae is a diverse, mostly tropical group. Extant representatives of Dryophthorini and Stromboscerini that live under the bark of trees were found in all ambers. Free-living forms are not noted. Representatives of Cossoninae are common in Eocene and Miocene ambers [2,111,112]. They are also common in Rovno and Baltic amber, which explains their relationship with wood. Cossonines are not found in Oise amber. The Conoderinae is very rare in amber. Several species of the genus Ceutorhynchus develop on herbaceous vegetation, and one species of the genus Baltocoeliodes in Baltic amber is probably associated with woody angiosperms. An undescribed species of the genus Ceutorhynchus was discovered in Rovno amber. Representative of Conoderini are only found in the earliest Eocene Oise amber. The Bariditae is absent in Eocene ambers, but it is one of the usual forms in the Early–Middle Eocene Green River deposits and terminal Eocene of the Florissant [113,114]. The Curculioninae is diverse only in Baltic amber, where species from Curculionini, Ellescini, Anthonomini, Eugnomini, Rhamphini, Mecinini, Tychiini, and Camarotini are represented. It can be noted that Eugnomini is now distributed in the Chilean–Patagonian and Australian regions, and Camarotini in the New World. The extinct genus of Acalyptini is described from Oise amber. Protoceletes hirtus belonging to this subfamily was found in Rovno amber. The largest modern Entiminae is relatively poorly represented in amber. Only one species from Eudiagogini is described from Oise amber. Several species from Naupactini and Anypotactini are known in Rovno amber. Baltic amber Entiminae is more diverse. Representatives of Hyperini, Sitonini, Tropiphorini, Anypotactini, Naupactini, Polydrusini, and Trachyphloeini also are found in Baltic amber. Species of the genus Paonaupactus are common Baltic amber weevils. The earliest Eocene Oise amber is very different from Late Eocene amber. None of the known genera in Oise amber are found in Baltic and Rovno ambers. The South African tribe Conoderini and the American extinct tribe Sciabregmini and Recent Eudiagogini are not found in Late Eocene amber. Rovno amber at the tribal level is a depleted version of Baltic amber. While all the genera, excluding Stenommatomorphus, are general, all species are different.
Figure 14.
Composition of the subfamilies of Curculionidae from Eocene amber.

The Platypodidae was recently discovered in Baltic amber. This is the extinct genus Eoplatypus with one species and the close American genus Cenocephalus with one species. The rarity of the Platypodidae fundamentally distinguishes the weevil faunas in Dominican and Baltic amber. Platypodidae is a massive group in Dominican amber [115].
The Scolytidae is a widely distributed group. Representatives of bark beetles from Oise amber are not described [20]. The fauna of Scolytidae of Rovno and Baltic amber is very different. Twenty-one species of bark beetles are described from Baltic amber. There are mainly species of the genera Hylurgops and Phloeosinus. Both of these genera are absent in Rovno amber. The Rovno amber genera, such as Xylechinus and Klesovia, were not found in Baltic amber. The genus Taphramites is common in both ambers, but it is represented by different species in Baltic and Rovno amber. Representatives of Hylesininae dominate in both ambers. The Scolytinae is represented by two genera of Dryocoetini.
To summarize, we can say that there are few similarities between the faunas of the earliest Eocene Oise amber and Late Eocene ambers. Rovno amber can be regarded as a depleted variant of Baltic amber, with no common species in their respective faunas.
Funding
The study was partially supported by the Russian Foundation for Basic Research (project Nos. 18-04-00243-a and 19-04-00465-a) and the Federal Fundamental Scientific Research Program for 2013–2020 (project no. AAAA-A16-116121410121-7).
Acknowledgments
The author thanks many colleagues for their assistance throughout his studies: V. I. Alekseev, A. R. Manukyan and A. V. Smirnova (Kaliningrad Regional Amber Museum, Russia: Kaliningrad), A. Allen (USA: Boise), V. M. Aniskin and N. A. Maslov (Khristianovich Institute of Theoretical and Applied Mechanics, SB RAS, Russia: Novosibirsk), A. Bukejs (Institute of Life Sciences and Technologies, Daugavpils University, Latvia: Daugavpils), L. Behne (Senckenberg Deutsches Entomologisches Institut, Germany: Müncheberg), D. Berthet (Centre de Conservation du musée des confluences, France, Lyon), J. Damzen (Lithuania: Vilnius), R. Yu. Dudko (Institute of Systematics and Ecology of Animals SB RAS, Russia: Novosibirsk), F. Eichmann (Germany: Hannover), A. Górski (Poland: Bielsko-Biaùa), C. Gröhn (Germany: Glinde), V. A. Gusakov (Russia: Moscow), O. Jaeger and K.-D. Klass (Museum für Tierkunde, Senckenberg Naturhistorische Sammlungen Dresden, Germany: Dresden), A. G. Kirejtshuk and B. A. Korotyaev (Zoological Institute RAS, Russia: St. Petersburg), U. Kotthoff (Center of Natural History, Germany: Hamburg), P. Limbourg (Institut royal des Sciences naturelles de Belgique, Belgium: Bruxelles), N.V. Martynovich (Museum of the World Ocean, Russia: Kaliningrad), M. A. Mazur (Department of Biosystematics, Opole University, Poland: Opole), A. Nel and H. Perrin (Muséum national d’histoire naturelle, France: Paris), N.B. Nikitsky (Zoological Museum, Moscow University, Russia: Moscow), E. E. Perkovsky and V. Y. Nazarenko (Schmalhausen Institute of Zoology, NASU, Ukraine: Kiev), A. G. Ponomarenko, A. P. Rasnitsyn, E. D. Lukashevich, D. E. Shcherbakov, I. D. Sukatsheva, D. V. Vassilenko (Borissiak Paleontological Institute RAS, Russia: Moscow), E. Yu. Shevnin (Russia: Novosibirsk), A. Yu. Solodovnicov and L. B. Vilhelmsen (Zoological Museum, University of Copenhagen, Denmark: Copenhagen), K. Szczepaniak (Earth Institute, Poland: Warsaw), J. Willers (Museum für Naturkunde, Leibniz-Institut für Evolutions-und Biodiversitätsforschung, Germany: Berlin), G. O. Poinar, Jr. (Oregon State University, USA: Corvallis). The four reviewers are also acknowledged for the valuable comments that improved the manuscript.
Conflicts of Interest
The author declares no conflicts of interest.
References
- Oberprieler, R.G.; Anderson, R.S.; Marvaldi, A.E. Curculionoidea Latreille, 1802: Introduction, Phylogeny. In Handbook of Zoology. Arthropoda: Insecta. Coleoptera, Beetles. Volume 3: Morphology and Systematics (Phytophaga); Leschen, R.A.B., Beutel, R.G., Eds.; Walter de Gruyter: Berlin, Germany; New York, NY, USA, 2014; pp. 285–300. [Google Scholar]
- Legalov, A.A. Fossil Mesozoic and Cenozoic weevils (Coleoptera, Obrienioidea, Curculionoidea). Paleontol. J. 2015, 49, 1442–1513. [Google Scholar] [CrossRef]
- Legalov, A.A. Annotated key to weevils of the world. Part 1. Families Nemonychidae, Anthribidae, Belidae, Ithyceridae, Rhynchitidae, Brachyceridae and Brentidae. Ukr. J. Ecol. 2018, 8, 780–831. [Google Scholar] [CrossRef]
- Thompson, R.T. Observations on the morphology and classification of weevils (Coleoptera, Curculionoidea) with a key to major groups. J. Nat. Hist. 1992, 26, 835–891. [Google Scholar] [CrossRef]
- Kuschel, G. A phylogenetic classification of Curculionoidea to families and subfamilies. Mem. Entomol. Soc. Wash. 1995, 14, 5–33. [Google Scholar]
- Alonso-Zarazaga, M.A.; Lyal, C.H.C. A World Catalogue of Families and Genera Curculionoidea (Insecta: Coleoptera) (Excepting Scolytidae and Platypodidae); Entomopraxis: Barcelona, Spain, 1999. [Google Scholar]
- Oberprieler, R.G.; Marvaldi, A.E.; Anderson, R.S. Weevils, weevils, weevils everywhere. Zootaxa 2007, 1668, 491–520. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Davis, S.R.; Shih, C.; Ren, D.; Pang, H. The earliest fossil record of Belidae and its implications for the early evolution of Curculionoidea (Coleoptera). J. Syst. Palaeontol. 2019, 17, 2105–2117. [Google Scholar] [CrossRef]
- Gratshev, V.G.; Legalov, A.A. New Mesozoic Ithyceridae beetles (Coleoptera). Paleontol. J. 2011, 45, 76–81. [Google Scholar] [CrossRef]
- Legalov, A.A. First record of Anthribid beetles from the Jurassic of Kazakhstan (Coleoptera: Anthribidae). Paleontol. J. 2011, 45, 629–633. [Google Scholar] [CrossRef]
- Legalov, A.A. Fossil history of Mesozoic weevils (Coleoptera: Curculionoidea). Insect Sci. 2012, 19, 683–698. [Google Scholar] [CrossRef]
- Gratshev, V.G.; Legalov, A.A. The Mesozoic stage of evolution of the family Nemonychidae (Coleoptera, Curculionoidea). Paleontol. J. 2014, 48, 851–944. [Google Scholar] [CrossRef]
- Zherichin, V.V.; Gratshev, V.G. Fossil Curculionid beetles (Coleoptera, Curculionoidea) from the Lower Cretaceous of Northeastern Brazil. Paleontol. J. 2004, 38, 528–537. [Google Scholar]
- Kirejtshuk, A.G.; Azar, D.; Beaver, R.A.; Mandelshtam, M.Y.; Nel, A. The most ancient bark beetle known: A new tribe, genus and species from Lebanese amber (Coleoptera, Curculionidae, Scolytinae). Syst. Entomol. 2009, 34, 101–112. [Google Scholar] [CrossRef]
- Legalov, A.A. The oldest Brentidae and Curculionidae (Coleoptera: Curculionoidea) from the Aptian of Bon-Tsagaan. Hist. Biol. 2014, 26, 6–15. [Google Scholar] [CrossRef]
- Legalov, A.A. A new weevil, Burmorhinus georgei gen. et sp. nov. (Coleoptera; Curculionidae) from the Cretaceous Burmese amber. Cretac. Res. 2018, 84, 13–17. [Google Scholar] [CrossRef]
- Legalov, A.A.; Nel, A.; Kirejtshuk, A.G. New and little known weevils (Coleoptera: Curculionoidea) from the Paleocene of Menat (France). Comptes Rendus Palevol 2017, 16, 248–256. [Google Scholar] [CrossRef]
- Kirejtshuk, A.G.; Nel, A. Current knowledge of Coleoptera (Insecta) from the lowermost Eocene Oise amber. Insect Syst. Evol. 2013, 44, 175–201. [Google Scholar] [CrossRef]
- Kirejtshuk, A.G.; Legalov, A.A.; Nel, A. A new genus of the subfamily Apioninae (Coleoptera: Brentidae) from the Lower Eocene Oise amber. Paleontol. J. 2015, 49, 1436–1441. [Google Scholar] [CrossRef]
- Legalov, A.A.; Kirejtshuk, A.G.; Nel, A. New weevils (Coleoptera, Curculionoidea) from the earlymost Eocene Oise amber. Paleontol. J. 2019, 53, 63–85. [Google Scholar] [CrossRef]
- Germar, E.F. Insekten in Bernstein eingeschlossen, beschrieben aus dem academischen Mineralien Cabinet zu Halle. Mag. Entomol. 1813, 1, 11–18. [Google Scholar]
- Motschulsky, V. Voyages. Lettres de M. de Motschulsky a M. Ménétriés, 4. Étud. Entomol. 1857, 5, 21–38. [Google Scholar]
- Schedl, K. Die Borkenkäfer des baltischen Bernsteins. Zentralb. Ges. Entomol. 1947, 2, 12–45. [Google Scholar]
- Voss, E. Einige Rhynchophoren der Bernsteinfauna (Col.). Mitt. Geol. Paläontol. Inst. Hambg. 1953, 22, 119–140. [Google Scholar]
- Voss, E. Einige Rüsselkäfer der Tertiärzeit aus baltischen Bernstein (Coleoptera, Curculionidea). Steenstupia 1972, 2, 167–181. [Google Scholar]
- Zherikhin, V.V. On weevils (Insecta, Coleoptera) from the Baltic amber. Tr. Paleontol. Inst. Akad. Nauk SSSR 1971, 130, 197–209. [Google Scholar]
- Wagner, H. Ein neues Apion aus dem baltischen Bernstein (Coleoptera, Curculionidae). Dtsch. Entomol. Z. 1924, 1924, 134–136. [Google Scholar]
- Hustache, A. Un curculionide de l’ambre de la Baltique. Bull. Mens. Soc. Lin. Lyon 1942, 11, 108–109. [Google Scholar] [CrossRef]
- Ulke, T. A new genus and species of Curculionidae (Coleoptera) in Baltic amber. Not. Nat. 1947, 19, 1–5. [Google Scholar]
- Wanat, M.; Borovec, L. New genus of weevil (Coleoptera, Curculionidae) from Baltic amber. Pol. Pismo Entomol. 1986, 56, 243–247. [Google Scholar]
- Kuska, A. Three new species of beetles (Coleoptera: Cantharidae, Anobiidae, Curculionidae) from the Baltic amber. Ann. Up. Sil. Mus. Bytom Entomol. 1992, 3, 107–113. [Google Scholar]
- Kuska, A. New beetle species (Coleoptera, Cantharidae, Curculionidae) from the Baltic amber. Prace Muz. Ziemi. 1996, 44, 13–23. [Google Scholar]
- Gratshev, V.G.; Zherikhin, V.V. A new anthribid genus from the Baltic amber (Insecta: Coleoptera: Anthribidae). Mitt. Geol. Paläontol. Inst. Hambg. 1995, 78, 149–157. [Google Scholar]
- Rheinheimer, J. Neue fossile Rüsselkäfer (Coleoptera: Curculionidae) aus dem Eozän des Baltischen Bernsteins und der Grube Messel bei Darmstadt. Staatl. Mus. Nat. 2007, 365, 1–24. [Google Scholar]
- Riedel, A. A new tribe, genus and species of Nemonychidae from Baltic amber (Coleoptera: Curculionoidea: Nemonychidae: Cimberidinae). Insect Syst. Evol. 2010, 41, 29–38. [Google Scholar] [CrossRef]
- Riedel, A.; dos Santos Rolo, T.; Cecilia, A.; van de Kamp, T. Sayrevilleinae Legalov, a newly recognised subfamily of fossil weevils (Coleoptera, Curculionoidea, Attelabidae) and the use of synchrotron microtomography to examine inclusions in amber. Zool. J. Linn. Soc. 2012, 165, 773–794. [Google Scholar] [CrossRef] [Green Version]
- Yunakov, N.N.; Kirejtshuk, A.G. New genus and species of broad-nosed weevils from Baltic amber and notes on fossils of the subfamily Entiminae (Coleoptera, Curculionidae). ZooKeys 2011, 160, 73–96. [Google Scholar] [CrossRef] [PubMed]
- Legalov, A.A. A new weevil genus of the tribe Metrioxenini (Coleoptera: Belidae) in Eocene Baltic amber. Hist. Biol. 2012, 24, 213–217. [Google Scholar] [CrossRef]
- Legalov, A.A. New Curculionoid beetles (Coleoptera: Curculionoidea) from the Baltic amber. Paleontol. J. 2012, 46, 262–272. [Google Scholar] [CrossRef]
- Legalov, A.A. New and little known Apioninae (Coleoptera, Brentidae) in Eocene Baltic amber. Evraz. Entomol. Zhurn. 2012, 11, 219–222. [Google Scholar]
- Legalov, A.A. New and little known weevils (Coleoptera: Curculionoidea) from the Paleogene and Neogene. Hist. Biol. 2013, 25, 59–80. [Google Scholar] [CrossRef]
- Legalov, A.A.; Bukejs, A. Succinapion telnovi n. gen. et n. sp. of the tribe Kalcapiini (Coleoptera: Brentidae: Apioninae) in Baltic amber. Hist. Biol. 2014, 26, 603–607. [Google Scholar] [CrossRef]
- Legalov, A.A.; Bukejs, A. A new species of the genus Archaeocallirhopalus (Coleoptera: Curculionidae) in Baltic amber. Entomol. Fenn. 2015, 26, 25–29. [Google Scholar]
- Legalov, A.A. Two new genera and four new species of fossil weevils (Coleoptera: Curculionoidea) in Baltic amber. Entomol. Fenn. 2016, 27, 57–69. [Google Scholar] [CrossRef] [Green Version]
- Legalov, A.A. New weevils (Curculionidae) in Baltic amber. Paleontol. J. 2016, 50, 970–985. [Google Scholar] [CrossRef]
- Legalov, A.A. Two new weevil genera of the family Brentidae (Coleoptera) in Baltic amber. Entomol. Fenn. 2018, 29, 161–168. [Google Scholar] [CrossRef] [Green Version]
- Legalov, A.A.; Bukejs, A. A new genus of the tribe Ceutorhynchini (Coleoptera: Curculionidae) in Baltic amber. Entomol. Fenn. 2018, 29, 185–190. [Google Scholar] [CrossRef] [Green Version]
- Legalov, A.A.; Bukejs, A. New findings of weevils (Coleoptera, Curculionioidea) in Baltic amber. Balt. J. Coleopterol. 2018, 18, 179–183. [Google Scholar]
- Bukejs, A.; Legalov, A.A. Groehnius, a new genus of Eugnomini (Coleoptera: Curculionidae) from Eocene Baltic amber. Foss. Rec. 2019, 22, 45–49. [Google Scholar] [CrossRef]
- Bukejs, A.; Legalov, A.A. A new species of the genus Dorytomus Germar, 1817 (Coleoptera, Curculionidae) from Baltic amber. Entomol. Fenn. 2019, 30, 173–178. [Google Scholar] [CrossRef] [Green Version]
- Legalov, A.A. A new genus of the tribe Curculionini (Coleoptera: Curculionidae) from Baltic amber. Paleontol. J. 2019, 53, 54–57. [Google Scholar] [CrossRef]
- Kania, J.; Legalov, A.A. A new genus of tooth-nosed snout weevils (Coleoptera: Rhynchitidae) in baltic amber. Paleontol. J. 2019, 53, 58–62. [Google Scholar] [CrossRef]
- Legalov, A.A.; Poinar, G.O., Jr. Two new species and new findings of Curculionoidea (Insecta: Coleoptera) from Baltic amber. Ukr. J. Ecol. 2019, in press. [Google Scholar]
- Bachofen-Echt, A. Der Berstein und Seine Einschlüsse; Springer: Wien, Austria, 1949. [Google Scholar]
- Larsson, S.G. Baltic Amber. A palaeobiological study. Entomonograph 1978, 1, 1–192. [Google Scholar]
- Alekseev, V.I. Coleoptera from the middle-upper Eocene European ambers: Generic composition, zoogeography and climatic implications. Zootaxa 2017, 4290, 401–443. [Google Scholar] [CrossRef]
- Berendt, G. Die im Bernstein befindlichen organischen Reste der Vorvelt; Nikolaische Buchhandlung: Berlin, Germany; pp. 1845–1856.
- Helm, O. Beitrage zur Kenntniss den Insekten des Bernstein. Schr. Naturforschenden Ges. Danz. N. F. 1896, 8, 220–231. [Google Scholar]
- Spahr, U. Systematischer Katalog der Bernstein- und Kopal-Kaefer (Coleoptera). Stuttg. Beitr. Naturkunde 1981, 80, 1–107. [Google Scholar]
- Alekseev, V.I. The beetles (Insecta: Coleoptera) of Baltic amber: The checklist of described species and preliminary analysis of biodiversity. Zool. Ecol. 2013, 23, 5–12. [Google Scholar] [CrossRef]
- Klebs, R. Über Bernsteinschlüsse im allgemein und die Coleopteren meiner Bernsteinsammlung. Schr. Phys.-Ökonom. Gesellsch. Königsberg Pr. 1910, 51, 217–242. [Google Scholar]
- Hieke, F.; Pietrzeniuk, E. Die Bernstein-Käfer des Museums für Naturkunde, Berlin (Insecta, Coleoptera). Mitt. Zool. Mus. Berl. 1984, 60, 297–326. [Google Scholar]
- Kubisz, D. Fossil beetles (Coleoptera) from Baltic amber in the collection of the Museum of Natural History of ISEA in Krakow. Pol. Pismo Entomol. 2000, 69, 225–230. [Google Scholar]
- Gratshev, V.G.; Perkovsky, E.E. New species of the genus Glaesotropis (Insecta: Coleoptera: Anthribidae) from Rovno amber. Paleontol. J. 2008, 42, 60–63. [Google Scholar]
- Petrov, A.V.; Perkovsky, E.E. New species of bark beetles from the Rovno amber (Insecta: Coleoptera: Scolytidae). Paleontol. J. 2008, 42, 406–408. [Google Scholar] [CrossRef]
- Nazarenko, V.Y.; Perkovsky, E.E. A new genus and species of Dryophthorid weevils (Coleoptera, Dryophthoridae: Stromboscerinae) from the Rovno amber. Paleontol. J. 2009, 43, 1097–1100. [Google Scholar] [CrossRef]
- Nazarenko, V.Y.; Legalov, A.A.; Perkovsky, E.E. A new species of the genus Caulophilus Woll. (Coleoptera: Curculionidae: Cossoninae) from the Rovno amber. Paleontol. J. 2011, 45, 287–290. [Google Scholar] [CrossRef]
- Nazarenko, V.Y.; Perkovsky, E.E. A new species of derelomine weevils (Coleoptera, Curculionidae, Curculioninae: Acalyptini) from the Rovno amber. Paleontol. J. 2016, 50, 991–996. [Google Scholar] [CrossRef]
- Petrov, A.V.; Perkovsky, E.E. A new genus and species of Scolytinae (Coleoptera: Curculionidae) from the Rovno amber. Paleontol. J. 2018, 52, 164–167. [Google Scholar] [CrossRef]
- Legalov, A.A.; Nazarenko, V.Y.; Perkovsky, E.E. A new genus of fungus weevils (Coleoptera: Anthribidae) in Rovno amber. Foss. Rec. 2018, 21, 207–212. [Google Scholar] [CrossRef] [Green Version]
- Bukejs, A.; Legalov, A.A. First record of the tribe Naupactini (Coleoptera: Curculionidae) in Rovno amber. Foss. Rec. 2019, 22, 25–30. [Google Scholar] [CrossRef] [Green Version]
- Legalov, A.A.; Nazarenko, V.Y.; Perkovsky, E.E. New weevils (Coleoptera: Curculionidae) from Rovno amber. Paleontol. J. 2019, 53, 63–77. [Google Scholar] [CrossRef]
- Bukejs, A.; Legalov, A.A. The first record of Rhynchitidae (Coleoptera) from Rovno amber. Entomol. Fenn. 2019, 30, 168–172. [Google Scholar] [CrossRef] [Green Version]
- Seyfullah, L.J.; Beimforde, C.; Dal Corso, J.; Perrichot, V.; Rikkinen, J.; Schmidt, A.R. Production and preservation of resins—Past and present. Biol. Rev. 2018, 93, 1684–1714. [Google Scholar] [CrossRef]
- Schubert, K. Neue Untersuchungen über Bau und Leben der Bernsteinkiefern (Pinus succinifera (Conw.) emend.). Beih. Geol. Jahrb. 1961, 45, 1–149. [Google Scholar]
- Thomas, B.R. Kauri resins—Modern and fossil. Org. Geochem. 1969, 599–618. [Google Scholar] [CrossRef]
- Grimaldi, D.J. Amber. Window to the Past; American Museum of Natural History; Abrams: New York, NY, USA, 1996. [Google Scholar]
- Weitschat, W.; Wichard, W. Baltic amber. In Biodiversity of Fossils in Amber from the Major World Deposits; Penney, D., Ed.; Siri Scientific Press: Manchester, UK, 2010; pp. 80–115. [Google Scholar]
- Sadowski, E.-M.; Schmidt, A.R.; Kunzmann, L.; Gröhn, C.; Seyfullah, L.J. Sciadopitys cladodes from Eocene Baltic amber. Bot. J. Linn. Soc. 2016, 180, 258–268. [Google Scholar] [CrossRef] [Green Version]
- Perkovsky, E.E.; Zosimovich, V.Y.; Vlaskin, A.P. Rovno Amber. In Biodiversity of Fossils in Amber from the Major World Deposits; Penney, D., Ed.; Siri Scientific Press: Manchester, UK, 2010; pp. 116–136. [Google Scholar]
- Mänd, K.; Muehlenbachs, K.; McKellar, R.C.; Wolfe, A.P.; Konhauser, K. Distinct origins for Rovno and Baltic ambers: Evidence from carbon and hydrogen stable isotopes. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2018, 505, 265–283. [Google Scholar] [CrossRef]
- Nel, A.; Brasero, N. Oise amber. In Biodiversity of Fossils in Amber from the Major World Deposits; Penney, D., Ed.; Siri Scientific Press: Manchester, UK, 2010; pp. 137–148. [Google Scholar]
- De Franceschi, D.; De Ploëg, G. Origine de l’ambre des facies sparnaciens (Eocene inferieur) du bassin de Paris: Le bois de l’arbre producteur. Geodiversitas 2003, 25, 633–647. [Google Scholar]
- Legalov, A.A. Phylogenetic reconstruction of weevils superfamily Curculionoidea (Coleoptera) using the SYNAP method. Biol. Bull. 2006, 33, 127–134. [Google Scholar] [CrossRef]
- Legalov, A.A. Annotated key to weevils of the world. Part 2. Subfamily Molytinae (Coleoptera, Curculionidae). Ukr. J. Ecol. 2018, 8, 340–350. [Google Scholar]
- Legalov, A.A. Annotated key to weevils of the world. Part 3. Subfamily Conoderinae (Coleoptera, Curculionidae). Ukr. J. Ecol. 2018, 8, 494–503. [Google Scholar]
- Legalov, A.A. Annotated key to weevils of the world. Part 4. Subfamilies Erirhininae, Dryophthorinae and Cossoninae (Curculionidae). Ukr. J. Ecol. 2019, in press. [Google Scholar]
- Legalov, A.A. Annotated key to weevils of the world. Part 5. Subfamily Entiminae (Curculionidae). Ukr. J. Ecol. 2019, in press. [Google Scholar]
- Lawrence, J.F.; Beutel, R.G.; Leschen, R.A.B.; Slipinsky, S.A. Chapter 2. Glossary of Morphological Terms. In Handbook of Zoology. Arthropoda: Insecta. Coleoptera, Beetles. Volume 2: Morphology and systematics (Elateroidea, Bostrichiformia, Cucujiformia partim); Kristensen, N.P., Beutel, R.G., Eds.; Walter de Gruyter: Berlin, Germany; New York, NY, USA, 2010; pp. 9–20. [Google Scholar]
- Kuschel, G. Reappraisal of the Baltic Amber Curculionoidea described by E. Voss. Mitt. Geol. Paläontol. Inst. Hambg. 1992, 73, 191–215. [Google Scholar]
- Pelsue, F.W.; O’Brien, C.W. A redefinition of the Curculionini of the world, with a key to subtribes and genera, and two new genera: Pseudoculio and Megaoculis (Coleoptera: Curculionidae: Curculioninae). Zootaxa 2011, 3102, 27–49. [Google Scholar] [CrossRef]
- Czeczott, H. The flora of the Baltic amber and its age. Prace Muz. Ziemi. 1961, 4, 119–145. [Google Scholar]
- Peris, D.; Solórzano Kraemer, M.M.; Smith, S.M.; Cognato, A.I. Eoplatypus jordali gen.n. et sp.n., the first described Platypodinae (Coleoptera: Curculionidae) from Baltic amber. Arthropod Syst. Phyl. 2017, 75, 85–194. [Google Scholar]
- Hagedorn, M. Borkenkäfer des baltischen Bersteins. Schr. Phys.-Ökonom. Gesell. König. Prus. 1906, 47, 115–121. [Google Scholar]
- Kuschel, G. Past and present of the relict family Nemonychidae (Coleoptera, Curculionoidea). GeoJournal 1983, 7, 499–504. [Google Scholar] [CrossRef]
- Legalov, A.A. Contribution to the knowledge of the family Nemonychidae (Coleoptera) with descriptions of new taxa. Ukr. J. Ecol. 2017, 7, 64–87. [Google Scholar] [CrossRef]
- Kuschel, G.; Poinar, G.O. Libanorhinus succinus gen. et sp. n. (Coleoptera: Nemonychidae). Entomol. Scand. 1993, 24, 143–146. [Google Scholar] [CrossRef]
- Peris, D.; Peris, D.; Davis, S.R.; Engel, M.S.; Delclos, X. An evolutionary history embedded in amber: Reflection of the Mesozoic shift in weevil-dominated (Coleoptera: Curculionoidea) faunas. Zool. J. Linn. Soc. 2014, 171, 534–553. [Google Scholar] [CrossRef]
- Legalov, A.A.; Azar, D.; Kirejtshuk, A.G. A new weevil (Coleoptera; Nemonychidae; Oropsini trib. nov.) from Lower Cretaceous Lebanese amber. Cretac. Res. 2017, 70, 111–116. [Google Scholar] [CrossRef]
- Poinar, G., Jr.; Brown, A.E.; Legalov, A.A. A new weevil, Aepyceratus hyperochus gen. et sp. nov., Aepyceratinae subfam. nov., (Coleoptera; Nemonychidae) in Burmese amber. Cretac. Res. 2017, 77, 75–78. [Google Scholar] [CrossRef]
- Kuschel, G. The Nearctic Nemonychidae (Coleoptera, Curculionoidea). Entomol. Scand. 1989, 20, 121–171. [Google Scholar] [CrossRef]
- Kuschel, G. The Palaearctic Nemonychidae (Coleoptera: Curculionoidea). Ann. Soc. Entomol. Fr. 1993, 29, 23–46. [Google Scholar]
- Rheinheimer, J. Illustrierter Katalog und Bibliographie der Anthribidae der Welt (Insecta: Coleoptera). Mitt. Entomol. Ver. Stuttg. 2004, 39, 1–288. [Google Scholar]
- Legalov, A.A. Annotated checklist of fossil and recent species of the family Belidae (Coleoptera) from the World Fauna. Amur. Zool. Zh. 2009, 1, 296–324. [Google Scholar]
- Marvaldi, A.E.; Lyal, C.H.C.; Oberprieler, R.G.; Bradbury, T.; Anderson, R.S. Phylogeny of the Oxycoryninae s. l. (Coleoptera: Belidae) and evolution of host-plant associations. Invert. Syst. 2006, 20, 447–476. [Google Scholar] [CrossRef]
- Legalov, A.A. Leaf-Rolling Weevils (Coleoptera: Rhynchitidae, Attelabidae) of the World Fauna; Agro-Siberia: Novosibirsk, Russia, 2007. [Google Scholar]
- Legalov, A.A. Revision der Gattung Auletes Schoenherr (Coleoptera, Rhynchitidae, Auletini). Bull. l’Institut R. Sci. Nat. Belg. Entomol. 2002, 72, 175–180. [Google Scholar]
- Tröster, G. Wasserkäfer und andere Raritäten—Neue Coleoptera-Funde aus den mitteleozänen Tonsteinen der Grube Messel bei Darmstadt. Kaupia 1993, 2, 145–154. [Google Scholar]
- Wappler, T. Die Insekten aus dem Mittel-Eozän des Eckfelder Maares, Vulkaneifel. Mainz. Naturwiss. Arch. 2003, 27, 1–234. [Google Scholar]
- Legalov, A.A.; Wappler, T. The oldest record of straight-snouted weevils (Coleoptera: Curculionoidea: Brentidae: Brentinae) from the Eocene of Germany. Hist. Biol. 2020. [Google Scholar] [CrossRef]
- Davis, S.R.; Engel, M.S. Cossonine weevils in Dominican amber (Coleoptera: Curculionidae). Linz. Biol. Beitr. 2007, 39, 803–820. [Google Scholar]
- Poinar, G., Jr.; Legalov, A.A. New species of the subfamily Cossoninae (Coleoptera: Curculionidae) in Dominican amber. Hist. Biol. 2015, 27, 491–502. [Google Scholar] [CrossRef]
- Scudder, S.H. Tertiary rhynchophorus Coleoptera of the United States. Monogr. US Geol. Surv. Tert. 1893, 21, 1–206. [Google Scholar] [CrossRef] [Green Version]
- Legalov, A.A. New weevils (Coleoptera, Curculionoidea) from the Eocene of the Green River, United States: Part 2. Paleontol. J. 2018, 52, 421–428. [Google Scholar] [CrossRef]
- Bright, D.E.; Poinar, G.O., Jr. Scolytidae and Platypodidae (Coleoptera) from Dominican amber. Ann. Entomol. Soc. Am. 1994, 87, 170–195. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
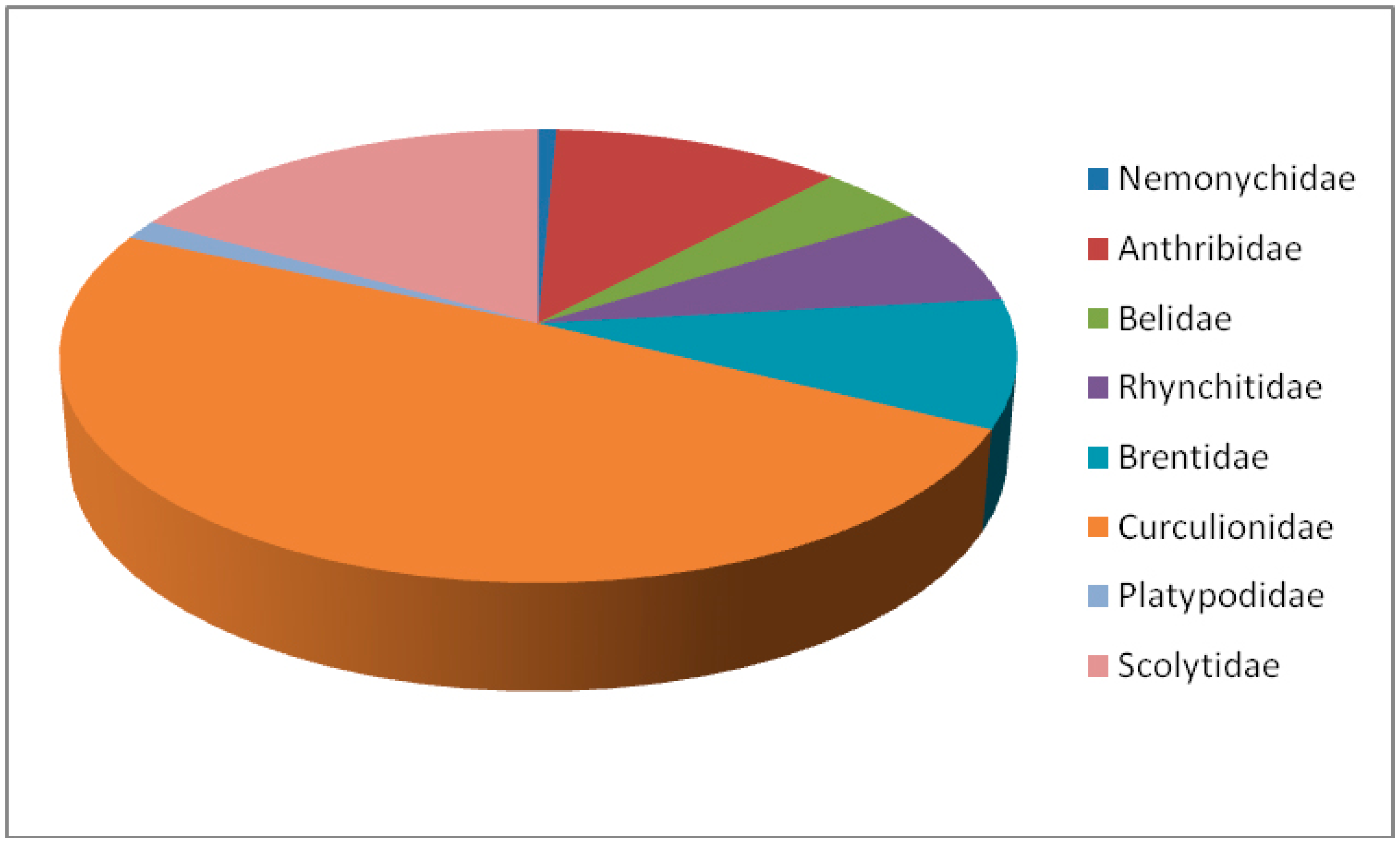{kind=link}
{kind=link}
Table 1.
List of Curculionoidea from Eocene ambers. BalJ—Baltic amber, OisJ—Oise amber, and RovJ—Rovno amber.
Table 1.
List of Curculionoidea from Eocene ambers. BalJ—Baltic amber, OisJ—Oise amber, and RovJ—Rovno amber.
| Family | Subfamily | Tribe | Genus | Species | Locality |
|---|---|---|---|---|---|
| Nemonychidae | Cimberidinae | Kuschelomacerini | Kuschelomacer Riedel, 2010 | K. kerneggeri Riedel, 2010 | BalJ |
| Anthribidae | Anthribinae | Oiserhinini | Oiserhinus Legalov, Kirejtshuk et Nel, 2019 | O. insolitus Legalov, Kirejtshuk et Nel, 2019 | OisJ |
| Mecocerini | Pseudomecocerusgen. nov. | P. alekseevisp. nov. | BalJ | ||
| Allandrini | Pseudomecorhis Voss, 1953 | P. orlovi Zherikhin, 1971 | BalJ | ||
| P. simulator Voss, 1953 | BalJ | ||||
| Allandroides Legalov, 2015 | A. vossi Legalov, 2015 | BalJ | |||
| Zygaenodini | Glaesotropis Gratshev et Zherikhin, 1995 | G. balticussp. nov. | BalJ | ||
| G. martynovi Legalov, 2012), comb. nov. | BalJ | ||||
| G. diadiasashai Gratshev et Perkovsky, 2008 | RovJ | ||||
| G. gusakovi Legalov, 2015 | BalJ | ||||
| G. minor Gratshev et Zherikhin, 1995 | BalJ | ||||
| G. succiniferus Legalov, 2015 | BalJ | ||||
| G. weitschati Gratshev et Zherikhin, 1995 | BalJ | ||||
| G. alleni Legalov, 2015 | BalJ | ||||
| G. gratshevi Legalov, 2015 | BalJ | ||||
| G. zherikhini (Legalov, 2013) | BalJ | ||||
| Choraginae | Valenfriesiini | Eduardoxenus Legalov, Nazarenko et Perkovsky, 2018 | E. unicus Legalov, Nazarenko et Perkovsky, 2018 | RovJ | |
| Belidae | Oxycoryninae | Oxycraspedini | Oxycraspedus Kuschel, 1955 | O. poinari Legalov, 2016 | BalJ |
| Metrioxenini | Archimetrioxena Voss, 1953 | A. electrica Voss, 1953 | BalJ | ||
| = Palaeometrioxena Legalov, 2012, syn. nov. | A. zherikhini (Legalov, 2012), comb. nov. | BalJ | |||
| Succinometrioxena Legalov, 2012 | S. attenuata Legalov et Poinar, in lit. | BalJ | |||
| S. bachofeni Legalov, 2013 | BalJ | ||||
| S. poinari Legalov, 2012 | BalJ | ||||
| Rhynchitidae | Sayrevilleinae | Sanyrevilleini | Baltocar Kuschel, 1992 | B. convexus Legalov, 2015 | BalJ |
| B. groehni Riedel, 2012 | BalJ | ||||
| B. hoffeinsorum Riedel, 2012 | BalJ | ||||
| B. subnudus Riedel, 2012 | BalJ | ||||
| B. succinicus (Voss, 1953) | BalJ | ||||
| Rhynchitinae | Auletini | Electrauletes Legalov, 2015 | E. unicus Legalov, 2015 | BalJ | |
| Eoropseudauletes Kania et Legalov, 2019 | E. plucinskii Kania et Legalov, 2019 | BalJ | |||
| Pseudomesauletes Legalov, 2001 | P. groehni Bukejs et Legalov, 2019 | RovJ | |||
| Rhynchitini | Eocenorhynchites Legalov, 2012 | E. vossi Legalov, 2012 | BalJ | ||
| Succinorhynchites Legalov, 2013 | S. alberti Legalov, 2013 | BalJ | |||
| Brentidae | Apioninae | Rhadinocybini | Baltocyba Legalov, 2018 | B. electrinus Legalov, 2018 | BalJ |
| Notapionini | Archinvolvulus Voss, 1972 | A. liquidus Voss, 1972 | BalJ | ||
| Palaeotanaini | Palaeotanaos Kirejtshuk, Legalov et Nel, 2015 | P. oisensis Kirejtshuk, Legalov et Nel, 2015 | OisJ | ||
| Aspidapiini | Pseudaspidapion Wanat, 1990 | P. khnzoriani (Zherikhin, 1971) | BalJ | ||
| Baltoapiongen. nov. | B. gusakovi (Legalov, 2015), comb. nov. | BalJ | |||
| B. subdiscedens (Voss, 1953), comb. nov. | BalJ | ||||
| Kalcapiini | Melanapion Wagner, 1930 | M. poinari Legalov, 2015 | BalJ | ||
| M. wanati Legalov, 2012 | BalJ | ||||
| Succinapion Legalov et Bukejs, 2014 | S. telnovi Legalov et Bukejs, 2014 | BalJ | |||
| Piezotrachelini | Conapium Motschulsky, 1866 | C. alleni Legalov, 2012 | BalJ | ||
| Baltoconapiumgen. nov. | B. anderseni (Voss, 1972), comb. nov. | BalJ | |||
| Electrapion Wagner, 1924 | E. kuntzeni (Wagner, 1924) | BalJ | |||
| Nanophyinae | Nanophyini | Baltonanophyes Legalov, 2018 | B. crassirostre Legalov, 2018 | BalJ | |
| Curculionidae | Erirhininae | Dorytomini | Dorytomus Germar, 1817 | D. bukejsisp. nov. | BalJ |
| D. electrinus Legalov, 2016 | BalJ | ||||
| D. groehni Bukejs et Legalov, 2019 | |||||
| D. korotyaevisp. nov. | BalJ | ||||
| D. nudus Legalov, 2016 | BalJ | ||||
| D. vlaskini Legalov, Nazarenko et Perkovsky, 2019 | BalJ | ||||
| Molytinae | Acicnemidini | Electrotribus Hustache, 1942 | E. henningseni (Voss, 1972), comb. nov. | BalJ | |
| = Paleopissodes Ulke, 1947 | E. theryi Hustache, 1942 | BalJ | |||
| = Anchorthorrhinus Voss, 1953 | = Paleopissodes weigangae Ulke, 1947, syn. nov. | ||||
| = Isalcidodes Voss, 1953 | = Anchorthorrhinus incertus Voss, 1953 | ||||
| = Protoceletes Rheinheimer, 2007 | = Isalcidodes macellus Voss, 1953 | ||||
| E. wolfschwenningerae (Rheinheimer, 2007) | BalJ | ||||
| E. rarussp. nov. | BalJ | ||||
| Plinthini | Leiosoma Stephens, 1829 | L. klebsi Legalov, 2016 | BalJ | ||
| Sciabregmini | Sciabregma Scudder, 1893 | S. squamosa Legalov, Kirejtshuk et Nel, 2019 | OisJ | ||
| Aedemonini | Electrorhinusgen. nov. | E. friedhelmisp. nov. | BalJ | ||
| Cryptorhynchini | Oisecalles Legalov, Kirejtshuk et Nel, 2019 | O. latosquamosus Legalov, Kirejtshuk et Nel, 2019 | OisJ | ||
| Succinacalles Zherikhin, 1971 | S. uniqus Zherikhin, 1971 | BalJ | |||
| Dryophthorinae | Dryophthorini | Rhinoporkus Legalov, Kirejtshuk et Nel, 2019 | Rh. gratiosus Legalov, Kirejtshuk et Nel, 2019 | OisJ | |
| Stromboscerini | Palaeodexipeus Legalov, 2016 | P. kirejtshuki Legalov, 2016 | BalJ | ||
| Rovnoslonik Legalov, Nazarenko et Perkovsky, 2019 | R. damzeni Legalov, Nazarenko et Perkovsky, 2019 | RovJ | |||
| Stenommatomorphus Nazarenko, 2009 | S. hexarthrus Nazarenko, 2009 | RovJ | |||
| Cossoninae | Dryotribini | Ampharthropelma Voss, 1972 | A. decipiens Voss, 1972 | BalJ | |
| Caulophilus Wollaston, 1854 | C. martynovae Legalov, Nazarenko et Perkovsky, 2019 | RovJ | |||
| C. rarus Legalov, 2016 | BalJ | ||||
| C. squamosus Legalov, 2016 | BalJ | ||||
| C. sucinopunctatus (Kuska, 1992) | BalJ | ||||
| C. zherikhini Nazarenko, Legalov et Perkovsky, 2011 | RovJ | ||||
| Necrodryophthorus Voss, 1953 | N. inquilinus Voss, 1953 | BalJ | |||
| Synommatodes Voss, 1953 | S. patruelis (Voss, 1953) | BalJ | |||
| Electrocossonusgen. nov. | E. kirejtshukisp. nov. | BalJ | |||
| Conoderinae | Ceutorhynchini | Ceutorhynchus Germar, 1824 | C. alekseevi Legalov, 2016 | BalJ | |
| C. electrinus Legalov, 2016 | BalJ | ||||
| C. succinus Legalov, 2013 | BalJ | ||||
| Baltocoeliodes Legalov et Bukejs, 2018 | B. sontagae Legalov et Bukejs, 2018 | BalJ | |||
| Conoderini | Jantarhinus Legalov, Kirejtshuk et Nel, 2019 | J. compressus Legalov, Kirejtshuk et Nel, 2019 | OisJ | ||
| Curculioninae | Acalyptini | Jantaronosik Legalov, Kirejtshuk et Nel, 2019 | J. nebulosus Legalov, Kirejtshuk et Nel, 2019 | OisJ | |
| Curculionini | Pseudoergania Legalov, 2019 | P. perkovskyi Legalov, 2019 | BalJ | ||
| Baltocurculiogen. nov. | B. manukyanisp. nov. | BalJ | |||
| Ellescini | Succinostyphlus Kuska, 1996 | S. mroczkowskii Kuska, 1996 | BalJ | ||
| = Electrotribus erectosquamata Rheinheimer, 2007, syn. nov. | BalJ | ||||
| Pachytychius Jekel, 1861 | P. eocenicus Legalov, 2016 | BalJ | |||
| Eugnomini | Archaeoeugnomus Legalov, 2016 | A. balticus Legalov, 2016 | BalJ | ||
| Anthonoeugnomusgen. nov. | A. barsevskisisp. nov. | BalJ | |||
| Mazurieugnomusgen. nov. | M. pilosussp. nov. | BalJ | |||
| Groehnius Bukejs et Legalov, 2019 | G. electrum Bukejs et Legalov, 2019 | BalJ | |||
| G. parvumsp. nov. | BalJ | ||||
| Rhamphini | Palaeorhamphus Legalov, 2016 | P. damzenisp. nov. | BalJ | ||
| P. eichmannisp. nov. | BalJ | ||||
| P. primitivus Legalov, 2016 | BalJ | ||||
| Orchestes Illiger, 1798 | O. tatjanae Legalov, 2016 | BalJ | |||
| Tachyerges Schoenherr, 1825 | T. hyperoche Legalov et Poinar, in lit. | BalJ | |||
| Tychiini | Eocenesibinia Legalov, 2015 | E. prussica Legalov, 2016 | BalJ | ||
| Camarotini | Paleodontopusgen. nov. | P. smirnovaesp. nov. | BalJ | ||
| Curculioninae incertae sedis | "Protoceletes" | P. hirtus Nazarenko et Perkovsky, 2016 | RovJ | ||
| Entiminae | Hyperini | Palaeophelypera Legalov, 2013 | P. kuscheli Legalov, 2013 | BalJ | |
| Tropiphorini | Limalophus Scudder, 1893 | L. poinarisp. nov. | BalJ | ||
| Scuccinalophus Legalov, 2016 | S. attenboroughi Legalov, 2016 | BalJ | |||
| Eudiagogini | Tolstonosik Legalov, Kirejtshuk et Nel, 2019 | T. oisensis Legalov, Kirejtshuk et Nel, 2019 | OisJ | ||
| Anypotactini | Paonaupactus Voss, 1953 | P. gracilis Legalov, Nazarenko et Perkovsky, 2019 | BalJ | ||
| = Pareustolus Voss, 1953 | P. katyae Legalov, Nazarenko and Perkovsky, 2019 | BalJ | |||
| =Protonaupactus Zherikhin, 1971, syn. nov. | P. microphthalmus (Zherikhin, 1971) | BalJ | |||
| = Sucinophyllobius Wanat et Borowiec, 1986 | P. sitonitoides Voss, 1953 | BalJ | |||
| = Polydrosus scheelei Voss, 1953 | |||||
| = Pyllobius cephalotes Voss, 1972 | |||||
| =Otiorhynchus pellucidipes Voss, 1972 | |||||
| P. sobrinus (Voss, 1972) | BalJ | ||||
| P. viridis (Wanat et Borowiec, 1986) | BalJ | ||||
| Naupactini | Arostropsis Yunakov et Kirejtshuk, 2011 | A. groehni Yunakov et Kirejtshuk, 2011 | BalJ | ||
| A. gusakovisp. nov. | BalJ | ||||
| A. perkovskyi Bukejs | RovJ | ||||
| et Legalov, 2019 | |||||
| Polydrusini | Polydrusus Germar, 1817 | P. archetypus Zherikhin, 1971 | BalJ | ||
| P. zherikhinisp. nov. | BalJ | ||||
| BalJ | |||||
| Archaeosciaphilus Legalov, 2012 | A. marshalli Legalov, 2012 | BalJ | |||
| Trachyphloeini | Archaeocallirhopalus Legalov, 2013 | A. alekseevi Legalov et Bukejs, 2015 | BalJ | ||
| A. larssoni Legalov, 2013 | BalJ | ||||
| Curculionidae incertae sedis | Thryogenosoma Voss, 1953 | Th. cariniger (Motschulsky, 1857) | BalJ | ||
| = Erirhinoides Motschulsky, 1857 non Blanchard, 1849 | |||||
| Platypodidae | Tesserocerinae | Tesserocerini | Eoplatypus Cognato et Smith, 2019 | E. jordali Peris, Solórzano Kraemer et Cognato, 2017 | BalJ |
| Cenocephalus Chapuis, 1865 | C. aniskinisp. nov. | BalJ | |||
| Scolytidae | Hylesininae | Hylastini | Hylastes Erichson, 1836 | H. aterites Schedl, 1947 | BalJ |
| Hylurgops LeConte, 1876 | H. corpulentus Schedl, 1947 | BalJ | |||
| = Hylesinites Germar, 1813 | H. dubius (Hagedorn, 1906) | BalJ | |||
| = Hylastites Hagedorn, 1906 | H. electrinus (Germar, 1813) | BalJ | |||
| = Myelophilites Hagedorn, 1907 | H. granulatus (Schedl, 1947) | BalJ | |||
| = Hylescierites Schedl, 1947 | H. pilosellus Schedl, 1947 | BalJ | |||
| H. schellwieni (Hagedorn, 1906) | BalJ | ||||
| H. tuberculatus Schedl, 1947 | BalJ | ||||
| Hylurgini | Xylechinus Chapuis, 1869 | X. mozolevskae Petrov et Perkovsky, 2008 | RovJ | ||
| Klesovia Petrov et Perkovsky, 2018 | K. pubescens Petrov et Perkovsky, 2018 | RovJ | |||
| Xylechinites Hagedorn, 1906 | X. anceps Hagedorn, 1906 | BalJ | |||
| Phloeosinini | Phloeosinus Chapius, 1869 | Ph. assimilis (Schedl, 1947) | BalJ | ||
| = Phloeosinites Hagedorn, 1906 | Ph. brunni (Hagedorn, 1906) | BalJ | |||
| Ph. regimontanus (Hagedorn, 1906) | BalJ | ||||
| Ph. rehi (Hagedorn, 1906) | BalJ | ||||
| Ph. robustus (Schedl, 1947) | BalJ | ||||
| Ph. sexspinosus (Schedl, 1947) | BalJ | ||||
| Ph. tuberculifer (Schedl, 1947) | BalJ | ||||
| Ph. wolffi (Schedl, 1947) | BalJ | ||||
| Polygraphini | Carphoborus Eichhoff, 1864 | C. keilbachi (Schedl, 1947) | BalJ | ||
| = Carphoborites Schedl, 1947 | C. posticus (Schedl, 1947) | BalJ | |||
| Scolytinae | Dryocoetini | Taphramites Schedl, 1947 | T. gnathotrichus Schedl, 1947 | BalJ | |
| T. rovnoensis Petrov et Perkovsky, 2008 | RovJ | ||||
| Taphrorychus Eichhoff, 1878 | T. immaturatus Schedl, 1947 | BalJ |
© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Legalov, A.A. A Review of the Curculionoidea (Coleoptera) from European Eocene Ambers. Geosciences 2020, 10, 16. https://doi.org/10.3390/geosciences10010016
AMA Style
Legalov AA. A Review of the Curculionoidea (Coleoptera) from European Eocene Ambers. Geosciences. 2020; 10(1):16. https://doi.org/10.3390/geosciences10010016
Chicago/Turabian StyleLegalov, Andrei A. 2020. "A Review of the Curculionoidea (Coleoptera) from European Eocene Ambers" Geosciences 10, no. 1: 16. https://doi.org/10.3390/geosciences10010016
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.